Il "concetto biologico" di specie è, forse, il più celebre tra tutti e lo dobbiamo al famoso biologo tedesco Ernst Mayr. In pratica, due specie diverse, incrociandosi tra loro, non dovrebbero essere in grado di produrre una prole fertile. Questa concezione ha a lungo rappresentato la pietra angolare della biologia evolutiva, definendo la specie come un gruppo di individui che possono riprodursi tra loro e generare prole fertile, ma che sono isolati riproduttivamente da altri gruppi. Tuttavia, la questione è parecchio più intricata di così, e le scoperte scientifiche degli ultimi anni hanno costretto a rivalutare il ruolo degli incroci tra specie in natura, specialmente quando si tratta di ibridi che riescono, inaspettatamente, a mantenere la propria fertilità.
Un ibrido è ciò che deriva da un incrocio tra due individui di due taxa diversi, cioè tra due esemplari di due specie, di due generi o di due varietà (o altro) differenti. Affinché un ibrido possa originarsi, bisogna che tra i due genitori esista una compatibilità genetica. Più i due soggetti sono distanti tassonomicamente e più è improbabile che generino un ibrido. L'ibridazione, in questo senso generale, include non solo gli incroci interspecifici, ma anche quelli tra popolazioni, razze o cultivar all'interno di una medesima specie, un significato particolarmente rilevante in agronomia. Storicamente, il valore attribuito all'ibridismo come fenomeno biologico ha subito cambiamenti con lo sviluppo della scienza ed è in stretta relazione col significato attribuito in vari tempi all'espressione biologica "specie".
Il Concetto di Specie, la Sterilità Ibrida e le Loro Eccezioni
La concezione classica, consolidata da figure come Georges Cuvier, riteneva che le specie organiche fossero stabili e che l'incrocio tra due specie reali e diverse non desse prodotto, o lo desse sterile. Al contrario, due varietà della medesima specie, anche se molto differenti nell'aspetto morfologico, avrebbero dato prodotti fecondi. Questa distinzione, sebbene pratica per gli zootecnici (che differenziano gli "ibridi" sterili come il mulo dai "meticci" fecondi tra razze diverse), non è sempre sufficiente per il sistematico. Per molto tempo, essere ibridi in un mondo di specie non è stato semplice, e la sterilità è stata vista come una barriera riproduttiva fondamentale.
L'ibridazione è comunque molto rara in natura per vari motivi. Spesso, la segregazione riproduttiva è garantita da "impedimenti meccanici" o barriere ecologiche. Ad esempio, l'utilizzo di habitat diversi da parte di due specie potenzialmente interfeconde è un buon modo per far sì che queste non s'incontrino e cadano vittime dei propri istinti. Esistono anche impedimenti fisiologici, come diverse stagioni riproduttive, o barriere etologiche legate a un'insuperabile indifferenza sessuale tra i due individui di specie diversa, che possono impedire l'incrocio. Se però queste barriere vengono superate, e la nuova vita riesce a nascere - cosa non sempre assicurata - essa andrà incontro a un nuovo problema: riprodursi a sua volta.
La Base Cromosomica della Sterilità
Nell'uomo, come negli altri organismi eucarioti, i nuclei delle cellule del corpo contengono tutti i cromosomi caratteristici della specie in duplice copia (2n). Le uniche cellule cui è permesso di sottrarsi a questa regola e di avere un numero di cromosomi "dimezzato" (n) sono i gameti, le cellule uovo e gli spermatozoi. Il perché è di facile intuizione: durante la meiosi, il processo di formazione dei gameti a partire da una cellula 2n, i cromosomi omologhi (che sono "fratelli" nel senso che contengono gli stessi geni in sequenza ma possono avere diverse varianti, o alleli) si appaiano e avviene uno scambio reciproco di pezzetti tra loro, il crossing-over. Questo processo è l'essenza della riproduzione sessuale e permette la creazione di nuove varianti di geni che potranno portare vantaggi o svantaggi al loro futuro portatore.
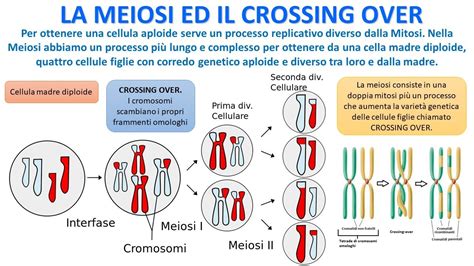
È proprio in questa fase cruciale che gli organismi ibridi figli di due specie con un numero di cromosomi diverso o con cromosomi strutturalmente incompatibili incontrano spesso problemi insormontabili. Gli ibridi sono di solito sterili in quanto incapaci di produrre spermatozoi e ovuli. La causa dipende principalmente da alterazioni che insorgono durante la gametogenesi, che ostacolano il corretto appaiamento dei cromosomi. Se i cromosomi dei due genitori non sono sufficientemente simili in numero o struttura, l'appaiamento e il crossing-over durante la meiosi vengono compromessi, portando alla formazione di gameti incompleti o non funzionali. Un esempio classico è il mulo, che possiede 63 cromosomi ed è il prodotto dell’incrocio tra una cavalla (64 cromosomi) e un asino (62 cromosomi). Questo numero dispari di cromosomi rende estremamente difficile il corretto appaiamento meiotico, risultando quasi sempre in sterilità per entrambi i sessi, sebbene con rare eccezioni di femmine fertili.
Queste problematiche mettono generalmente la parola fine agli incroci che sono riusciti a eludere i controlli precedenti, consegnando all'ibridazione un ruolo marginale nelle popolazioni naturali. Tuttavia, oggi sappiamo che circa il 10% degli animali ibridano almeno con un'altra specie. Questo non vuol dire che Mayr fosse solo un tizio con un interessante taglio di capelli, ma piuttosto che i casi scoperti negli ultimi anni hanno costretto a rivalutare il ruolo degli incroci tra specie in natura, portando a considerare la fertilità ibrida come un fenomeno, seppur raro, con significative implicazioni evolutive.
Quali sono le cause della sterilità femminile? Diagnosi e terapie.
Amissia: Le Barriere alla Fertilità Ibrida
L'osservazione che in molti casi l'unione è sterile e che in altri il prodotto di un'unione fertile ora è sempre sterile, ora è qualche volta fecondo, mentre talaltra è sempre fecondo, ha indotto i biologi di questo secolo a cercare le cause di queste differenze o per lo meno a stabilire quale sia il rapporto tra la fisiologia e la morfologia delle cellule germinali degli ibridi. La fecondità completa coincide con l'affinità gametica, ossia con la capacità delle cellule germinali a produrre germi che sviluppano normalmente tutte le parti di un organismo, comprese le ghiandole germinali stesse ed i loro prodotti; la sterilità coincide con l'amissia, ossia con l'attributo opposto all'affinità gametica.
L'amissia può essere di diverse tipologie:
- Amissia Primaria (o gametica reale): È data dall'incapacità di due cellule germinali di sesso differente a coniugarsi e a produrre un germe. Sperimentalmente è stata osservata l'esistenza di germi ibridi fra specie molto differenti, i quali periscono poco dopo l'inizio della segmentazione dell'uovo. In altri casi, lo sviluppo embrionale si arresta a stadi diversi; alcune dànno origine a larve che muoiono appena schiuse o prima di compiere la metamorfosi o in altri periodi dello sviluppo. Fin qui si potrebbe ancora parlare di amissia assoluta, per la mancata vitalità del prodotto.
- Amissia Secondaria: Questa coesiste con l'affinità gametica, quando è dovuta a cause secondarie, somatiche o di ambiente, che non riguardano la capacità di formazione del germe da parte di due cellule germinali prodotte da forme differenti, anche se esse sono soltanto razze della medesima specie. Basti qui ricordare i principali tipi:
- Amissia Psichica: Dovuta a una ripugnanza invincibile che una specie ha verso un'altra, come il caso dei bovini domestici e dei bufali, che non si incrociano.
- Amissia Etica: Derivante da costumi diversi. Esistono alcune specie di pesci molto affini tra loro che possono, con accorgimenti artificiali, mescolarsi e dare origine a una discendenza feconda, ma normalmente i loro prodotti germinali non possono coniugarsi, perché emessi in diversa stagione o in ambiente diverso, come la superficie di un lago o il suo fondo.
- Amissia Meccanica: Dovuta all'impossibilità di penetrazione di spermi in uova provviste di membrana più resistente di quella che appartiene alla specie donde derivano i primi o rivestite di guscio munito di apertura (micropilo) di diametro inferiore alle dimensioni dello spermio stesso. In questi ultimi casi ciò che non è possibile in un determinato senso è invece possibile nell'altro: il fatto è stato sperimentato in alcuni ricci di mare, dove gli spermi di Strongylocentrotus lividus fecondano facilmente le uova di Echinus microtuberculatus, ma non è possibile fecondare uova di Strongylocentrotus lividus con spermi di Echinus microtuberculatus per la ragione sopra indicata. Analogamente nei vegetali: Brassica oleracea non è fecondabile col polline di Brassica napus, mentre riesce l'incrocio inverso. Così pure sono fecondi i prodotti di Brassica sativa con B. oleracea, di Triticum vulgare con Aegilops ovata, di Mirabilis longiflora con M. jalapa.
Quando gli Ibridi Diventano Fertili: Il Fascino dell'Ereditarietà
Successivamente all'amissia primaria e ai casi di ibridi vitali ma infecondi, esiste una categoria di ibridi vitali e somaticamente vigorosi. Questi appartengono talvolta al medesimo sesso e sono sempre infecondi, anche con le specie progenitrici. Citiamo, per esempio, gli ibridi del pavone e del gallo con la gallina di Faraone, che sono sempre maschi e completamente sterili. L'affinità gametica di due specie diverse si inizia in forma saltuaria e accidentale, quando quelle generano ibridi normalmente sterili fra i quali tuttavia si osservano molto raramente individui fecondi in uno solo dei due sessi, individui capaci pertanto di procreare solo quando abbiano la possibilità di reincrociarsi con una delle specie pure dalle quali sono derivati.
In questa via si procede fino alla normale fecondità di uno dei due sessi, mentre l'altro rimane sterile: nelle piante e nei mammiferi la fecondità appartiene alle femmine ibride (gonomonoteliidia), mentre negli uccelli è limitata al sesso maschile (gonomonarrenia). Esistono inoltre specie le quali producono ibridi normalmente fecondi in uno dei due sessi, e raramente nell'altro.
Sotto questo aspetto, il Poll ha distinto gli ibridi in "toconti" (da "parto" e "bastardo"), che offrono la completa maturazione delle cellule germinali producendo spermatozoi e uova mature, e "steironoti" (da "sterile" e "bastardo"). Tuttavia, la completa maturità delle cellule germinali non è un indice assoluto e sicuro di fecondità, giacché esistono toconoti fecondi e toconoti sterili, indicando che la fecondità dipende anche da fattori specifici e individuali. Perché un ibrido raggiunga lo stato "trimitotico" (cioè con gametogenesi normale che include le tre divisioni mitotiche che portano alla maturazione dei gameti), bisogna che le due specie progenitrici abbiano apparati cromosomici e corredi fattoriali sufficientemente simili. Negli ibridi steironoti, si distinguono diversi gradi di incompletezza nella gametogenesi, dall'assenza totale di mitosi all'arresto a stadi intermedi.

Secondo la regola di Haldane, formulata raccogliendo numerosi dati statistici, quando in una specie di ibrido uno dei due sessi manca o è raro o è sterile, questo è sempre il sesso eterozigotico, mentre il sesso presente o comune o fecondo è quello omozigotico. Abbiamo visto infatti che negli ibridi di Uccelli sono spesso fecondi i maschi, il cui sesso è omozigotico (ZZ), mentre nei Mammiferi, nei quali è omozigotico il sesso femminile (XX), sono spesso feconde le femmine, e i maschi ibridi corrispondenti (XY) sono sterili. Così accade nei rari casi di fecondità delle mule e nella normale fecondità delle femmine ibride fra toro e bisonte e viceversa, fra toro e yak e viceversa. Analogamente nelle piante, i bastardi di Antirrhinum siculum × A. maius, di Triticum sativum × Secale cereale, di Triticum sativum × Aegilops ovata sono sterili nel sesso maschile e fecondi in quello femminile.
L'Aspetto Corporeo e il Vigore Ibrido
L'aspetto corporeo degli ibridi è generalmente intermedio a quello dei genitori, tanto se si considerano le caratteristiche morfologiche, quanto se si studia il loro comportamento fisiologico. Negli incroci tra le razze e le varietà di una medesima specie di animali domestici o di piante coltivate, i prodotti seguono le leggi dell'eredità alternata. Bisogna premettere che queste ultime sono generalmente differenti per caratteri dovuti a un grande numero di fattori ereditari.
Un fenomeno di grande importanza è l'eterosi, o vigore ibrido. Si ottiene per incrocio di due linee genetiche (generazione parentale; P) selezionate per capacità produttiva e abilità ricombinante e altamente omozigoti, essendo state auto-fecondate per più generazioni. Tale circostanza determina un forte indebolimento, noto come depressione da in-breeding, a cui segue, dopo l’incrocio delle due linee, la manifestazione di un intenso vigore ibrido, limitato alla generazione filiale F1, con forte sviluppo e incremento produttivo delle piante nate dai semi prodotti dall’incrocio. La depressione da in-breeding e il successivo vigore ibrido sono più marcati nelle piante allogame (abitualmente eterozigoti), che in piante autogame (omozigoti). In queste prime generazioni, gli ibridi sono solitamente più vigorosi, soprattutto se fertili, dopo l'incrocio. Infatti, più diversità genetica c'è tra i due genitori e più vigorose sono le piante (ma anche gli animali!). Il fenomeno è noto come hybrid vigor ed il suo contrario è la inbreeding depression.
Al fine di poter effettuare l’incrocio delle due linee autogame parentali (P), selezionate per produrre l’ibrido eterotico, si procede, in piante con fiori ermafroditi (es. riso, o frumento) alla loro emasculazione, con metodo meccanico, chimico, o genetico, per evitare che le piante di ciascuna linea autogama si auto-fecondino e quindi per ottenere solo semente derivata da tale incrocio. In piante i cui fiori maschili siano molto vicini a quelli femminili, i primi non possono essere tolti senza danneggiare quelli femminili, pertanto si preferisce emasculare col metodo della maschio-sterilità genetico-citoplasmatica, la quale dipende da geni presenti nei mitocondri degli individui maschio-sterili, che interagiscono con geni nucleari, omozigoti recessivi per il gene restauratore della fertilità.

Nei casi di ibridi fertili si ha, nelle generazioni successiva a F1, una crescente diminuzione del vigore ibrido, per vari motivi, tra cui si ritiene principalmente vi sia l’aumentare dell’eterozigosi e quindi segregazione di un crescente numero d’individui omozigoti per caratteri recessivi negativi. L’unico modo per rendere permanente il vigore ibrido della generazione F1 e tentare di ottenere semi geneticamente uguali alla pianta che li produce, sfruttando il fenomeno dell’agamospermia, una forma di apomissia, che consiste nella produzione di embrioni e quindi semi, senza missia (o gamia), ossia senza unione del gamete maschile e di quello femminile. L’agamospermia si verifica a volte in natura in vari generi di angiosperme, tra cui parenti selvatici di specie domesticate, ma il tentativo di ottenere varietà del tutto apomittiche è finora fallito. In senso generale, l'apomissia è la produzione di un clone d'individui derivati da un solo genitore e quindi include anche la riproduzione vegetativa.
Nuove Frontiere della Fertilità Ibrida: La Ricerca Genetica
Più nel dettaglio, nelle prove condotte su cellule uovo non fecondate i ricercatori avevano riscontrato che attivando un primo gene, il ‘Baby BooM 1’ (BBM1) iniziava lo sviluppo embrionale con produzione di semi agamospermi, sebbene solo nel 30% dei casi: il solo gene BBM1 risultava insufficiente a garantire un adeguato successo allo sviluppo embrionale. Come ha affermato il prof. Sundaresan: ‘Accendere artificialmente BBM1 nella cellula uovo è sufficiente per avviare l'embriogenesi e creare una nuova pianta (un nuovo seme), ma questo processo ha funzionato solo circa un terzo delle volte’. Successivamente, con la collaborazione di colleghi dell’Innovative Genomics Institute dell’Università della California, di Berkeley, i ricercatori hanno controllato quali geni fossero attivi in cellule uovo fecondate ed hanno in tal modo accentrato l’attenzione su un secondo gene, il ‘Wuschel-like HomeODomain 9’ (WOX9A) che, quando attivato in cellule uovo fecondate insieme al gene BBM1, incrementava al 90% (86%-91%) il tasso di successo dello sviluppo dell’embrione del seme. Il prof. Sundaresan ha così commentato questo risultato: ‘Pensiamo che BBM1 stia attivando un interruttore che prepara la cellula uovo alla transizione in embrione, ma l'interruttore non è fisso’.
In tal senso, come afferma Imtiyaz Khanday, autore corrispondente e assistente al Dipartimento di Scienza delle Piante del Collegio CAES dell’Università di Davis: ‘Gli aploidi sono strumenti preziosi nella selezione delle piante per la produzione di linee pure, che consentono una produzione uniforme del raccolto’. L’attivazione dello sviluppo embrionale di gameti diploidi, indotta dai geni BBM1 e WOX9A agli ibridi eterotici di prima generazione (F1) permetterebbe di stabilizzarne il vigore ibrido in quelle successive, risultando eliminate le ricombinazioni, che si verificano nella meiosi e quindi la variabilità. Tale complessiva procedura, studiata in questo caso sul riso, potrebbe essere applicata anche ad altre colture, in cui si utilizzino ibridi eterotici. Tutto ciò è più sinteticamente espresso dalle parole del prof. Sandarasan: ‘Se combiniamo questo trucco di trasformare una cellula uovo in un embrione senza fecondazione con un'altra tecnica che elimina la meiosi, possiamo produrre in modo efficiente semi ibridi ad alto rendimento’.
Ibridi Fertili: Un Percorso verso Nuove Specie
Da un ibrido tra due specie, se la progenie è stabile, si può anche generare una nuova specie, descrivibile come tutte le altre attraverso la pubblicazione di un articolo scientifico, anche nel caso in cui l’ibrido sia artificiale. Questa specie di origine ibrida può talvolta venire denominata con un segno "x" tra il nome del genere e l’epiteto specifico. Se un ibrido tra due specie è fertile può esserlo perché i cromosomi sono riusciti comunque a riformarsi o perché si sono uniti due gameti che insolitamente non abbiano fatto la meiosi, cioè non si è unito metà DNA del padre e metà DNA della madre, ma tutto il DNA del padre e tutto il DNA della madre.
I casi di ibridi fertili che danno origine a nuove specie sono particolarmente affascinanti. Tra questi, gli allotetraploidi, ormai non più in grado di reincrociarsi nemmeno con le specie che li hanno originati perché il loro numero di cromosomi è troppo diverso, sono un mezzo di speciazione relativamente comune. Le piante ad un livello più alto di ploidia possono avere caratteristiche diverse dai loro progenitori diploidi in quanto possono essere più resistenti in alcuni ambienti e risultare quindi avvantaggiate. Tipicamente, più è alto il numero di cromosomi e più una specie è mediamente resistente al freddo.
Ibridi nel Regno Animale: Storie di Incroci e Uniformità Cariologica
Gli animali ibridi rappresentano la prole prodotta dall’incrocio tra popolazioni geneticamente diverse. Essi differiscono dai genitori in termini di capacità riproduttiva, in quanto sono raramente fertili, a causa del numero dispari di cromosomi. Di solito, i maschi sono sempre sterili.
I pesci sono un esempio rilevante di ibridazione naturale, con frequenti ibridi della carpa e della tinca col carassio comune e col carassio dorato, dell'Abramis brama con la scardola e col leucisco, dell'alborella con la scardola, di alcune specie di trote, come pure dei coregoni fra loro. In tutti questi salmonidi, la facilità con la quale si compie la fecondazione artificiale favorisce la produzione degli ibridi. Nella classe degli Anfibî è noto l'ibrido naturale Triton Blasii che deriva dall'incrocio di Triton marmoratus con Triton cristatus.

Tra i cetacei, uno dei motivi del successo degli ibridi è quella che viene chiamata uniformità cariologica. I tursiopi (Tursiops truncatus), oltre a mangiare caramelle gommose, si spassano con le pseudorche (Pseudorca crassidens) e originano uno degli ibridi con il nome più interessante: il wholpin (whale+dolphin, da noi sarebbe balfino). Nel 1986, al largo delle coste islandesi venne catturata una balenottera. L’animale, una femmina, presentava caratteri intermedi tra una balenottera comune (Balaenoptera physalus) e una balenottera azzurra (Balaenoptera musculus). Analisi morfologiche e molecolari sul cadavere confermarono definitivamente che il cetaceo era, davvero, figlio dell’unione tra i due animali più grandi del pianeta. Il piccolo era ottenuto dall’accoppiamento con un maschio di B. musculus.
Un caso particolarmente studiato è quello della Stenella clymene, un delfino presente normalmente nell’Oceano Atlantico lungo tutta la fascia tropicale. Questo cetaceo ha sempre rappresentato un problema per la classificazione, in quanto presenta caratteristiche fisiche e comportamentali simili ad altri due membri del genere Stenella: S. attenuata e S. coeruleoalba. Ana Rita Amaral dell’Università di Lisbona, cercando di fare chiarezza, ha studiato la Stenella clymene e, analizzando il DNA nucleare e quello mitocondriale (mtDNA) - che, a differenza di quello nucleare, viene trasmesso unicamente per via materna ed è utile per ricostruire la parentela - ha proposto che S. clymene sia un ibrido, il "figlio del peccato", tra S. attenuata e S. coeruleoalba. Tuttavia, per "fissare" una nuova specie occorre che i suoi appartenenti si riproducano principalmente tra loro e, nel caso di S. clymene, la cosa non deve essere stata immediata. Tutte e tre le specie di Stenella vivono nella stessa zona e occupano nicchie ecologiche simili, quindi una segregazione sulla base di habitat o risorse usate non erano la risposta più semplice. L’ipotesi avanzata da Amaral e colleghi è che S. clymene si sia separata dalle altre due specie grazie a ciò che le aveva permesso di esistere in un primo momento: il sesso con gli individui giusti. È possibile che, al momento della ricerca di un partner, gli esemplari di S. clymene abbiano preferito individui visivamente e dal comportamento simili a loro (e quindi ibridi a loro volta) rispetto alle altre stenelle.
Tra gli esempi di ibridi animali sterili troviamo, oltre al mulo e al bardotto, lo yakalo, che è il frutto dell’accoppiamento di uno yak (Bos grunniens) e di una femmina di bisonte americano (Bison bison). Lo yakalo ha 60 cromosomi e, mentre i maschi sono sterili, le femmine risultano fertili. Dall’incrocio tra due grandi felini, il leone e la tigre, sono nati due noti ibridi, il tigon e il liger. Il tigon deriva da una tigre maschio e una leonessa, mentre il liger ha per madre una tigre e per padre un leone. Hanno entrambi 38 cromosomi e, come molti ibridi, i maschi di entrambe le specie tendono ad essere sterili, mentre le femmine possono essere fertili. Questi ibridi sono rari e solitamente si trovano solo in cattività, dato che le aree geografiche naturali di leoni e tigri non si sovrappongono.
Altri esempi di ibridi possono essere l'orso grolare (incrocio tra l'orso polare e il grizzly), il leopone (incrocio tra leone e leopardo, più grande e con una specie di criniera che parte dalla nuca) e il pumapardo (ibrido tra il puma e il leopardo). Tra gli uccelli, si conoscono ibridi dell'oca con l'anatra, del pavone con la gallina di Faraone, di questa col gallo, del gallo con la fagiana, del colombo con la tortora. Il Tetrao intermedius è un ibrido naturale tra il Gallo cedrone (Tetrao urogallus) e il Fagiano di monte (Lyrurus tetrix).
L'Ibridazione nel Regno Vegetale: Nuove Varietà e Poliploidia
In botanica, si utilizzano i processi di ibridazione al fine di modificare alcuni caratteri, farne emergere di nuovi, costituire nuove varietà. I fiori femminili vengono impollinati esclusivamente col polline della varietà selezionata ricorrendo ad accorgimenti che impediscano l'impollinazione da parte di altre piante (nelle specie dioiche) o dalla stessa (nelle specie monoiche). Per arrivare ad una nuova varietà o specie attraverso l'ibridazione è necessario rendere stabili i caratteri per mezzo di incroci successivi con l'obiettivo di ottenere individui omozigoti rispetto a quegli stessi caratteri.

Gli ibridi più comuni sperimentalmente sono quelli derivanti da incroci fra parecchie specie di fagiani, come il Dorato (Chrysolophus pictus) e quello di Lady Amherst (C. amherstiae), di Anatre, come Daphila acuta ed Anas boscas (Codone e Germano reale), di Fringillidi, come lucarino (Chrysomitris spinus) e canarino (Serinus canarius) fra gli Uccelli. Tra i Mammiferi invece gli incroci del cane col lupo e con lo sciacallo, quelli di alcune specie di pecore fra loro, di alcuni Bovidi, ecc. Tra gli ibridi "dimitotici" (che mostrano un grado intermedio di gametogenesi) vanno ascritti il comune incrocio fra l'Anatra comune (Anas boscas) e l'Anatra muta (Cairina moschata), gli ibridi tra vari generi di anatre e di fagiani, il mulo e il bardotto, gli zebroidi, i maschi di parecchi incroci fra Bovidi come toro e bisonte, toro e yak.
Le orchidee rappresentano un campo particolarmente ricco di esempi di ibridazione, sia naturale che artificiale. Nelle orchidee, la ploidia, ovvero il numero di "serie" di cromosomi presenti in ogni cellula, è un fattore cruciale. Le piante diploidi (2n) sono la norma e hanno due serie di cromosomi, mentre quelle riproduttive (gameti) hanno una sola serie (n). La fertilità degli ibridi di orchidee dipende spesso da quanti set di cromosomi riescono a combinare. Ad esempio, gli ibridi triploidi (3n) sono generalmente sterili, dato che hanno tre serie di cromosomi che non riescono ad appaiarsi correttamente durante la meiosi. Gli ibridi tetraploidi (4n), invece, avendo quattro serie di cromosomi, sono spesso fertili, dato che ogni cromosoma può trovare un omologo con cui appaiarsi. Questi possono incrociarsi con un gran numero di altre specie o ibridi.
L'ibridazione può produrre ibridi di prima generazione (F1) che, se riseminati, non daranno risultati omogenei, ma una variata seconda generazione (F2), anche se ci sarà stata impollinazione tra piante del solito ibrido o addirittura anche nel caso di autoimpollinazione. Quando invece sono vere F1 si possono comunque "disibridare", selezionando, nelle successive generazioni, solo le piante che presentano le stesse caratteristiche delle originali F1 fino a quando nasceranno solo piante con quelle caratteristiche.
Tra gli ibridi fertili, alcuni partecipano all'evoluzione in vario modo attraverso l'introgressione tra due specie. L'ibridazione interspecifica è il processo per cui due animali di specie diverse si riproducono dando vita a una progenie con caratteristiche genetiche (e spesso, morfologiche) intermedie rispetto alle specie parentali. Tale progenie può essere sterile o fertile. In quest'ultimo caso, spesso, gli ibridi sono capaci di reincrociarsi con una o entrambe le specie parentali ("backcrossing"), costituendo un vettore per il passaggio di geni tra una specie e l'altra (introgressione).
L'Ibridazione: Forza Evolutiva e Sfida di Conservazione
L'"ibridazione introgressiva" così risultante da un lato è una potente forza evolutiva, introducendo nuovi caratteri in popolazioni e specie, che possono o meno venire selezionati dall'ambiente; dall'altro, in alcuni casi, i nuovi caratteri introdotti in una specie possono risultare svantaggiosi e portare ad una riduzione della fitness e un aumento del rischio di estinzione. Tali considerazioni si estendono anche a popolazioni della stessa specie con caratteristiche differenti (ad esempio, "razze" diverse).
Gli ibridi possono nascere fondamentalmente in due modi: per ibridazione naturale o per ibridazione antropogenica. L'ibridazione naturale consiste nell’accoppiamento naturale tra individui di due distinte specie, i cui geni si mescolano nella prole, causando cambiamenti sia nel suo genotipo, cioè nella sua composizione genetica, che nel suo fenotipo, cioè nelle sue caratteristiche fisiche e comportamentali. In altre parole, i figli di questi accoppiamenti sono dotati di una combinazione unica di tratti, che ereditano dai genitori di specie diverse.
L’ibridazione antropogenica è invece il risultato di azioni provocate dall’uomo, direttamente da parte degli allevatori, o indirettamente, ad esempio attraverso l’introduzione, volontaria o non deliberata, di nuove specie in nuovi ambienti o attraverso la frammentazione dell'habitat, che è a propria volta causata da processi che portano le specie animali a migrare o fuggire, come l'aumento della velocità e dell'intensità della deforestazione, la desertificazione, e l'urbanizzazione. Un caso particolarmente "problematico" riguarda l'ibridazione tra conspecifici domestici e selvatici (es. cinghiale e maiale, cane e lupo, ecc…). Poiché le pressioni selettive che agiscono sulle popolazioni in cattività sono molto diverse da quelle che agiscono sulle popolazioni selvatiche (rilassamento della pressione selettiva naturale, forte selezione artificiale), molto spesso i caratteri degli animali domestici, che possono passare alle popolazioni selvatiche tramite ibridazione introgressiva, risultano fortemente svantaggiosi (es. riduzione della fitness). L'ibridazione artificiale è diventata particolarmente attraente per gli allevatori, grazie alle elevate caratteristiche produttive della generazione F1 di ibridi, che rappresenta la prima generazione di discendenti diretti. Questi ibridi sono molto resistenti alle malattie e crescono tanto e in fretta.

In conclusione, sebbene il concetto biologico di specie di Mayr abbia fornito una solida base per la comprensione della diversità biologica, la realtà dell'ibridazione, in particolare quella che porta a prole fertile, continua a sfidare e ad arricchire la nostra visione dell'evoluzione. La complessa interazione dei cromosomi durante la meiosi, le diverse forme di amissia, e i meccanismi genetici che possono superare la sterilità, rivelano un paesaggio biologico dinamico in cui i confini tra le specie sono spesso più porosi di quanto si credesse, offrendo vie inaspettate per l'emergere di nuove forme di vita.