Lo sviluppo di un nuovo organismo rappresenta un percorso di straordinaria complessità, caratterizzato da una precisa sequenza di eventi biologici che, a partire da una singola cellula fecondata, danno origine a strutture sempre più elaborate. In questo viaggio primordiale, la formazione di cavità e membrane extraembrionali riveste un ruolo cruciale, fornendo protezione, nutrimento e un ambiente ideale per la crescita. Questo articolo esplora le tappe fondamentali di questo sviluppo, dall'aggregato iniziale di cellule noto come morula, attraverso la formazione delle complesse strutture annesse all'embrione come la cavità amniotica, fino a considerare la profonda risonanza di questi spazi interni nel contesto dell'esistenza umana.
Il Concepimento: L'Inizio di Ogni Vita
Come gli altri animali, l'uomo garantisce la continuità della sua specie nel tempo e la trasmissione delle sue caratteristiche alle generazioni future attraverso il processo della riproduzione, che consiste in una precisa sequenza di eventi biologici molto complessi. Il primo atto di tale processo è la fecondazione, ovvero l'unione tra due gameti, uno maschile, lo spermatozoo, e l'altro femminile, l'ovocita. I gameti sono cellule altamente specializzate, caratterizzate dal possedere nel proprio nucleo un corredo di 23 cromosomi (numero aploide), a differenza di tutte le altre cellule somatiche della specie umana, che ne hanno 46 (numero diploide).
L'interazione e la successiva fusione dei due gameti avviene a livello della porzione ampollare della tuba, cioè quella più prossima all'ovaio. Per poter fecondare l'ovocita, gli spermatozoi debbono essere precedentemente attivati, un processo definito 'capacitazione', che si verifica all'interno dei genitali femminili, avendo inizio in vagina per poi proseguire nel restante tratto genitale femminile e soprattutto nella cervice uterina. La capacitazione comprende un insieme di cambiamenti nello spermatozoo, come riarrangiamenti della struttura della membrana plasmatica, incremento della sua permeabilità agli ioni calcio mediato da una proteina chiamata calmodulina, attivazione di un meccanismo di produzione energetica catalizzato dall'enzima adenil-ciclasi e riduzione della carica elettrica negativa. È secondaria alla rimozione di fattori 'decapacitanti' dalla superficie esterna dei gameti maschili, presenti nel plasma seminale.
Tappa finale del processo di capacitazione è la 'reazione acrosomiale', che indica una serie di modificazioni a carico della struttura della membrana dello spermatozoo, e che si realizza a livello degli spazi compresi tra le cellule che circondano l'ovocita (dette del cumulo ooforo) e la zona esterna, o 'pellucida', dell'ovocita. Alla zona pellucida aderiscono, in un primo momento, più spermatozoi; questa prima fase è aspecifica e reversibile. Dopo un breve intervallo si realizza la seconda fase, in cui si stabilisce un legame molto tenace e irreversibile tra i due gameti. Lo spermatozoo penetra nell'ovocita attraverso la zona pellucida, percorrendo un itinerario non radiale, impostogli dalla particolare disposizione della struttura microfibrillare di questa, costituita da mucopolisaccaridi e da glicoproteine. La penetrazione è agevolata dalla motilità della testa e della coda dello spermatozoo, conseguenza delle ultime tappe del processo di capacitazione, e dalle contrazioni che si verificano nell'ovocita, rese possibili dall'elevata presenza in questo di proteine filamentose contenenti actinomiosina.
Dopo aver attraversato la zona pellucida, lo spermatozoo penetra all'interno di uno spazio situato tra questa e la membrana plasmatica dell'ovocita, lo 'spazio perivitellino'. Talora tale spazio viene raggiunto da vari spermatozoi, che per tale motivo vengono definiti 'supplementari', ma in condizioni di normalità, dopo l'attraversamento di uno spermatozoo, la zona pellucida diviene impermeabile ad altri. Infatti, come diretta conseguenza del processo di attraversamento, si verifica un massiccio e rapido rilascio di ioni calcio dai depositi intracellulari, che a sua volta pare innescare uno scambio tra ioni sodio e idrogenioni con un conseguente incremento del pH intracellulare; a seguito di tale incremento alcuni elementi costituenti l'ovocita modificano le loro caratteristiche biochimiche e biofisiche. Il potenziale elettrico di membrana raggiunge, in pochi secondi, valori compresi tra -10 mV e +20 mV, contro i +60 mV di partenza. Queste modificazioni, che rendono la membrana impenetrabile ad altri spermatozoi, vanno sotto il nome di 'reazione stromale'.
La penetrazione dello spermatozoo porta, come diretta conseguenza, l'attivazione dell'ovocita, cioè il completamento della seconda divisione meiotica del gamete femminile, con formazione del pronucleo femminile. Il nucleo dello spermatozoo, che è riuscito a entrare nel citoplasma dell'ovocita (ooplasma), rapidamente si rigonfia, il suo materiale nucleare cromatinico si decondensa e si assiste alla rottura della membrana nucleare, con dispersione della cromatina in essa contenuta nel circostante citoplasma; in un secondo tempo la cromatina viene circondata da una serie di vescicole che, fondendosi tra loro, creano una membrana bilaminare e formano il pronucleo maschile. Nell'ooplasma si assiste alla organizzazione di un importante sistema di microtubuli e di microfilamenti (costituiti da fibrina), che si mostra essenziale non soltanto nel permettere il completamento della meiosi dell'ovocita stesso, ma anche nella migrazione dei due pronuclei che vengono a diretto contatto tra loro al centro dell'ooplasma. Infatti, alcuni microtubuli si dispongono tra i pronuclei, condensandosi sulla superficie delle loro membrane, che vengono successivamente disintegrate. La fusione dei due pronuclei dà origine a una nuova entità cellulare diploide, lo zigote.
Come nasce un essere umano: l'incredibile percorso dalla fecondazione al parto
La Morula: Il Primo Aggregato Cellulare Compattato
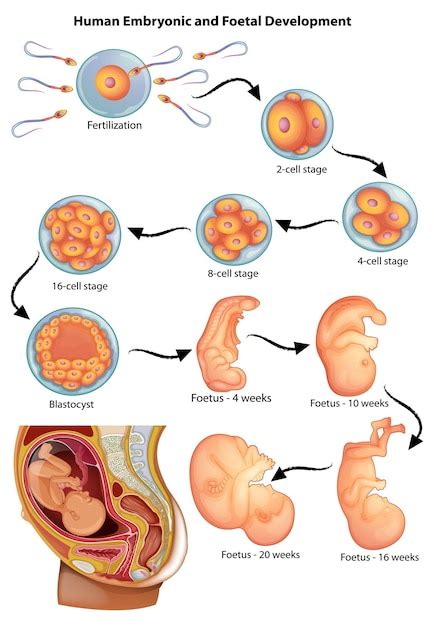
Dopo circa 30 ore dalla fecondazione, l'embrione appare costituito già da due cellule, chiamate 'blastomeri', essendosi verificata la prima di una serie successiva di divisioni mitotiche, che si realizzano nello stadio detto della 'segmentazione' o del 'clivaggio'. Per queste divisioni, i blastomeri diventano quattro, poi otto, e così di seguito. Nell'uomo, come negli altri mammiferi, tale fase procede in modo lento, con un intervallo di circa 14-26 ore tra i differenti cicli di duplicazione cellulare. Questi eventi si realizzano nel lume tubarico e in tale fase la tuba stessa provvede, con il suo fluido, a sostenere il metabolismo e il conseguente sviluppo embrionale.
Le cellule ottenute da ogni nuova divisione mitotica sono di dimensioni ridotte, poiché decresce progressivamente il rapporto citoplasma-nucleo; in tal modo, al crescere del numero delle cellule, il volume totale dell'embrione non muta e può essere facilmente contenuto all'interno della zona pellucida. La morula è la fase che attraversa un organismo durante i primi stadi della gestazione. È composta da un aggregato di cellule che si formano nei primi stadi dello sviluppo embrionale, durante la segmentazione dello zigote. Questo processo di segmentazione porta, al terzo giorno, alla formazione della morula, che corrisponde allo stadio di 8-16 blastomeri.
Durante il terzo giorno, i blastomeri si compattano non lasciando spazi tra loro, e iniziano un processo di profonda modifica della struttura cellulare: aumenta la sintesi proteica, avviene la polarizzazione delle cellule interne, mentre quelle periferiche restano apolari e diventano più piccole di quelle interne, formando tra loro giunzioni occludenti. Nella morula si possono distinguere due gruppi di cellule: i blastomeri situati più perifericamente, che successivamente daranno luogo al trofoblasto, e quelli situati più centralmente che, invece, daranno luogo all'embrioblasto, o embrione vero e proprio. Quindi, anche se allo stato di morula i blastomeri sembrano uguali, in realtà, sono già destinati a compiti differenti. Le cellule interne sono invece unite da giunzioni comunicanti. Sulle zone in contatto con l'esterno delle membrane plasmatiche si formano inoltre microvilli. Allo stadio di morula, tutte le cellule sono ancora cellule staminali "totipotenti", anche se dopo un paio di divisioni diventano pluripotenti, cioè indifferenziate e in grado di originare tutti i tessuti embrionali ed extra-embrionali. La morula è ancora circondata dalla zona pellucida.
Dalla Morula alla Blastocisti e l'Annidamento Uterino
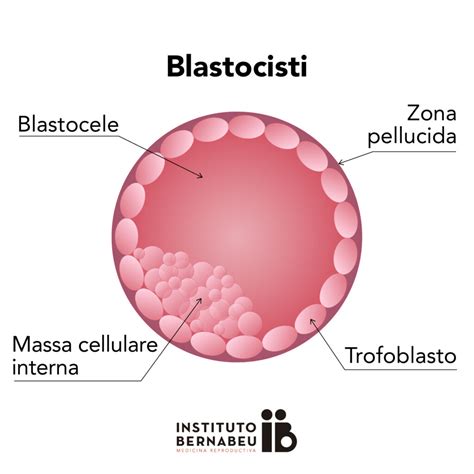
Lo stadio di morula di norma viene raggiunto al compimento del terzo giorno dalla fecondazione. In tale arco di tempo, l'embrione si sposta dall'ampolla tubarica per raggiungere la cavità uterina. Le conoscenze attualmente ottenute, grazie agli studi compiuti sulla fecondazione in vitro, fanno ritenere che la morula raggiunga la cavità uterina allo stadio di 12-16 blastomeri. Giunta nell'utero, la morula subisce alcune modificazioni strutturali. Al quarto giorno, il germe entra nella cavità uterina e iniziano a comparire tra le cellule spazi pieni di fluido che si uniscono in uno solo, il blastocele. Questo avviene perché in essa, attraverso la zona pellucida, penetra del liquido che si dispone negli interstizi tra i blastomeri situati nella porzione centrale. All'aumentare della filtrazione di questo liquido, gli spazi tra i blastomeri si fondono e, confluendo, vanno a formare una cavità unica, che viene detta 'blastocele'.
A tale stadio, che solitamente viene raggiunto alla fine del quarto giorno, non si parla più di morula ma di 'blastocisti', e la zona pellucida che delimitava la morula è completamente scomparsa. La blastocisti permane libera nella cavità uterina per un periodo di circa tre giorni, quindi, grazie a periodiche contrazioni ed espansioni, assume una posizione adeguata all'impianto, un fenomeno definito dell'orientamento. Nella blastocisti, i due gruppi di blastomeri appaiono ormai facilmente differenziabili tra quelli centrali dell'embrioblasto, uniti tra loro e disposti a un polo della blastocisti detto 'polo animale', e quelli periferici del trofoblasto, che assumono una forma appiattita e vengono a costituire la parete stessa della blastocisti, anche a livello dell'embrioblasto che rivestono. Il trofoblasto è la prima struttura extraembrionale che si differenzia quando l’uovo fecondato, segmentandosi, ha assunto la forma di una piccola vescicola o blastocisti (quarto-quinto giorno dal momento della fecondazione). Esso comprende lo strato di cellule che costituisce la parete della blastocisti.
Verso il sesto giorno dalla fecondazione, le cellule del trofoblasto prendono contatto con le cellule dell'endometrio tramite sottili microvilli, in quello che è lo stadio dell'apposizione o primo stadio dell'impianto, e aderiscono tenacemente tra di loro. I microvilli vengono poi progressivamente sostituiti da piccole digitazioni irregolari e contemporaneamente le cellule epiteliali dell'endometrio, attraverso un 'bordo' ondulato prodotto dalla loro membrana plasmatica, si schiacciano contro le cellule del trofoblasto, questa è la fase dell'adesione o seconda fase dell'impianto. Si assiste infine alla terza e ultima fase dell'impianto, quella della migrazione-invasione, che si realizza tramite la fusione delle superfici cellulari adiacenti, ovvero cellule epiteliali endometriali da un lato e quelle del trofoblasto dall'altro. La penetrazione avviene nel settimo giorno attraverso enzimi secreti dalle cellule del trofoblasto che si trovano a diretto contatto con l'endometrio. Le propaggini trofoblastiche si insinuano tra le cellule epiteliali, fagocitandole, e approfondendosi progressivamente tra le cellule della mucosa uterina e poi nel sottostante stroma. Sembra che l'impianto si realizzi grazie all'azione svolta da alcuni enzimi proteolitici contenuti dalle cellule del trofoblasto, che venendo a contatto con le cellule endometriali ne determinerebbero l'erosione. Tale azione proteolitica della blastocisti sarebbe parallelamente promossa dalle cellule epiteliali dell'endometrio.
Al compimento dell'ottavo giorno dalla fecondazione, la blastocisti appare in parte inclusa nello stroma endometriale. Al di sopra dell'embrioblasto, ovvero in corrispondenza del polo embrionale, le cellule del trofoblasto appaiono disposte a formare un disco solido, costituito da due strati sovrapposti denominati rispettivamente 'citotrofoblasto' e 'sinciziotrofoblasto'. Il citotrofoblasto, più interno, è costituito da cellule mononucleate, ovvero munite di un solo nucleo, mentre il sinciziotrofoblasto, disposto esternamente, è costituito da cellule plurinucleate. Le cellule trofoblastiche si trasformano in sinciziotrofoblasto, che è formato da cellule polinucleate con attività invadente, dirompente, digerente. La sua attività serve a distruggere le cellule endometrali per far penetrare la blastocisti nell'endometrio e al contempo liberare nutrienti, fornendo così nutrizione istotrofica. Il sinciziotrofoblasto è responsabile della reazione deciduale e produce hCG, un fattore che salva il corpo luteo, trasformandolo in corpo luteo gravidico, impedendo quindi le mestruazioni. Nella prima settimana è necessaria la presenza di LH e progesterone in quantità maggiore rispetto a quella di estrogeni.
Contemporaneamente, la mucosa endometriale subisce una serie di modificazioni che la rendono idonea a ricevere la blastocisti: essa appare ispessita, edematosa, di colorito pallido, con ghiandole in intensa attività secretiva che riversano un secreto ricco di mucina e glicogeno. Le arterie che irrorano lo strato compatto e spugnoso della mucosa presentano un decorso tortuoso e realizzano un fitto letto capillare situato al di sotto della superficie dell'utero stesso. La blastocisti si impianta a livello della parete anteriore della cavità uterina, sul fondo, in una porzione dell'utero definita antimesomerica, corrispondente al punto di fusione delle due emicavità cornuali che durante lo sviluppo embrionale danno origine all'utero.
Le Strutture Extraembrionali e il Loro Ruolo Essenziale
Strutture che si sviluppano dall’uovo fecondato, parallelamente allo sviluppo dell’embrione, hanno la funzione di proteggere e di nutrire l’embrione stesso.
Il Trofoblasto e il Corion
In seguito, il trofoblasto prolifera e si differenzia nei due strati cellulari menzionati: uno profondo (citotrofoblasto) ed uno superficiale (sinciziotrofoblasto). Quest’ultimo si accresce particolarmente in corrispondenza del polo embrionale della blastocisti, in quella parte cioè che è adiacente all’embrione in via di sviluppo. Quando al di sotto del trofoblasto si dispone uno strato di cellule del mesoderma extraembrionale, la membrana che circonda l’embrione viene denominata corion. Il corion risulta formato da due componenti: il trofoblasto esternamente, ed il mesoderma extraembrionale internamente.
Sulla superficie esterna del corion si sviluppano, a partire dalla terza settimana circa, delle escrescenze ramificate denominate villi coriali, che si accrescono nella parte di corion corrispondente alla decidua basale, che è la parte profonda della mucosa uterina ove è avvenuto l’annidamento della blastocisti, interposta tra la blastocisti stessa e la muscolatura dell’utero.
La Placenta: Ponte Vitale tra Madre e Feto
In questa sede, la membrana coriale con i suoi villi e la decidua assumono rapporti molto stretti e costituiscono un organo di forma discoidale, la placenta. Questo organo provvede agli scambi di gas respiratori, di sostanze nutritizie, di prodotti di escrezione, tra l’organismo fetale e quello materno. Nella placenta, le parti di mucosa uterina interposte tra i vasi sanguigni materni e la superficie dei villi coriali regrediscono, per cui i villi stessi pescano direttamente in ampie lacune nelle quali circola il sangue materno. Nelle lacune si crea una rete di canali in cui il sangue derivante dai capillari materni arteriosi e venosi inizia a circolare (passaggio a nutrizione emotrofica), che si fonde tra loro a partire dal dodicesimo giorno. In particolare, nel sinciziotrofoblasto iniziano a crearsi delle lacune piene di sangue e fluidi ghiandolari. Al decimo giorno la blastocisti è già completamente penetrata nello stroma dell'endometrio, formando un tappo di chiusura costituito da un coagulo di sangue.
L'Amnios e la Cavità Amniotica: Un Ambiente Protetto e Fluido
Oltre al corion, un’altra membrana che avvolge l’embrione è l’amnios. Essa delimita una cavità, la cavità amniotica, piena di liquido nel quale l’organismo che si sta sviluppando risulta come sospeso. L’amnios comprende quella parte della parete della vescicola amniotica primitiva che non partecipa alla costituzione del disco embrionale, e si continua con l’ectoderma dell’embrione.
Compaiono vescicole tra le cellule del sinciziotrofoblasto e quelle della massa cellulare interna che confluiscono in una unica, che è la cavità dell'amnios, già nella prima settimana. In seguito, la cavità amniotica si ingrandisce notevolmente, e quindi l’amnios viene a contatto con la superficie interna del corion, fino al formarsi di un’unica membrana, la membrana corioamniotica. Inoltre, man mano che il corpo dell’embrione viene delimitato e sporge entro la cavità amniotica, l’amnios va a rivestire il peduncolo che collega il corpo dell’embrione alla membrana coriale e che contiene vasi sanguigni. Bonnie B. sottolinea che posteriormente, la cavità amniotica inizia a espandersi tutt’intorno all’embrione e quest’ultimo comincia a nuotare nel fluido amniotico.
Il Funicolo Ombelicale e le Vie di Comunicazione Essenziali
Questo peduncolo allungato, che rimane quindi compreso nella cavità amniotica, costituisce il funicolo ombelicale, e risulterà infine inserito sulla membrana coriale laddove si è formata la placenta. Il funicolo ombelicale costituisce quindi la struttura attraverso la quale il sangue fetale può circolare fino alla placenta, dove esso può ossigenarsi, assumere sostanze nutritizie dal sangue materno ed eliminare prodotti di rifiuto. La sua formazione è preceduta dalla comparsa di cavità nel mesoderma extraembrionale che durante il tredicesimo giorno confluiscono in una sola, chiamato celoma extraembrionale. Questo celoma circonda completamente il sacco vitellino e riduce il mesoderma extraembrionale a due foglietti: quello che si accolla al citotrofoblasto e all'amnios (somatopleura extraembrionale) e quello che si accolla al sacco vitellino, all'epiblasto e all'ipoblasto (splancnopleura extraembrionale). Essi si continuano l'uno nell'altro tramite il peduncolo di connessione, che diverrà il futuro cordone ombelicale.
Altri Annessi Embrionali: Il Sacco Vitellino e l'Allantoide
Nel processo di delimitazione del corpo dell’embrione si viene a formare un altro annesso embrionale, la vescicola ombelicale, che corrisponde alla parte del sacco vitellino secondario che non rimane inclusa nel corpo dell’embrione per formare l’intestino primitivo. La vescicola ombelicale dapprima comunica ampiamente con l’intestino primitivo. In seguito viene compresa nel funicolo ombelicale e si appiattisce contro la membrana coriale. Per un certo tempo comunica con l’intestino per mezzo di un condotto, il dotto enterovitellino, che percorre il funicolo ombelicale e che in seguito si oblitera. Il sacco vitellino è importante soprattutto perché nella sua parete si differenziano i primi isolotti di cellule del sangue.
Un’altra struttura annessiale è l’allantoide. Questa si forma come un diverticolo nella parte caudale del sacco vitellino quando il corpo dell’embrione viene ad essere delimitato dalle pieghe cefalica, caudale e laterali, e si peduncolizza. L’allantoide rimane in parte compresa entro il corpo dell’embrione, ove formerà la vescicola e l’uraco. La sua porzione extraembrionale rimane compresa nel funicolo ombelicale, e regredisce quasi completamente nel secondo mese di vita intrauterina.
Sviluppo Embrionale Avanzato: Dal Disco Bilaminare all'Organogenesi Complessa
Clinicamente, si è soliti dividere la gravidanza in tre trimestri. In embriologia, invece, lo sviluppo intrauterino viene suddiviso in due grossi periodi: quello embrionale, che va dalla quarta sino alla decima settimana di sviluppo, e quello fetale, dalla decima settimana sino al termine della gestazione. Nel primo periodo si formano gli abbozzi di tutti gli organi e apparati, nel secondo questi vanno incontro alla crescita e alla maturazione.
La Formazione del Disco Germinativo Bilaminare e Trilaminare
Mentre avviene l'impianto della blastocisti nell'endometrio, anche le cellule disposte nel polo embrionale, o embrioblasti, subiscono una serie di modificazioni, disponendosi in modo da formare due distinti strati e differenziandosi anche a livello strutturale. In particolare, alcune cellule, di piccola taglia e di forma cubica, si dispongono in unico strato a formare il cosiddetto foglietto germinativo entodermico, mentre i restanti embrioblasti, di forma cilindrica e di dimensioni maggiori rispetto alle precedenti, si dispongono sempre in un unico strato, a formare il foglietto germinativo ectodermico. Tali foglietti appaiono sovrapposti a formare un'unica struttura discoidale, nota come 'disco germinativo bilaminare'. In tale stadio gli embrioblasti non solo appaiono modificati nelle dimensioni e nella forma, ma soprattutto sono differenziati a seconda del foglietto d'appartenenza, così come saranno diversi i successivi processi di differenziazione e di modificazione.
Nel contesto del trofoblasto, si realizzano notevoli cambiamenti: esso appare ispessito, con uno spessore maggiore in corrispondenza del polo embrionale, dove compaiono dei veri e propri vacuoli, che aumentando progressivamente di dimensioni confluiscono in modo da formare delle lacune, donde la denominazione di 'stadio lacunare' per tale fase di sviluppo del trofoblasto. Nello stesso lasso di tempo il polo embrionale continua il suo incessante rimaneggiamento. Alcune cellule di forma appiattita si distaccano dal citotrofoblasto e formano una membrana molto sottile, la membrana di Heuser, che risulta essere in continuità con i bordi del foglietto entodermico, con il quale concorre a delimitare, rivestendola, la cavità esocelomatica o 'sacco vitellino primitivo'. Un'altra cavità, quella amniotica, rivestita dall'amnios, si forma fra l'ectoderma e il trofoblasto.
Verso il quindicesimo giorno, alcune cellule migrano dalla periferia verso l'asse centrale, creando un ispessimento chiamato linea primitiva. All'estremità anteriore della linea si forma un ispessimento chiamato nodo di Hensen. Quelle che si invaginano per prime si inseriscono tra epiblasto e ipoblasto, espandendosi e sostituendo man mano l'ipoblasto. Quelle che si invaginano successivamente si spostano lungo la membrana basale dell'epiblasto, riempiendo lo spazio tra epiblasto ed endoderma; questo è il mesoderma. Tutto questo processo prende il nome di gastrulazione, ed è accompagnato sia da un accrescimento del disco che da un allungamento, nonché dallo spostamento delle invaginazioni e del nodo di Hensen verso le zone posteriori, che verrà completato solo alla quarta settimana.
La Neurulazione e la Morfogenesi dell'Embrione
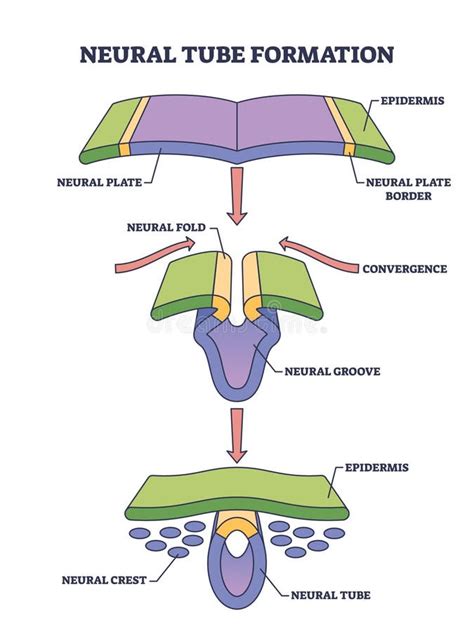
Al sedicesimo giorno si creano delle depressioni circolari dove l'ectoderma si salda fortemente con l'endoderma, una anteriormente e una posteriormente. Quella anteriore viene chiamata membrana faringea e la posteriore membrana cloacale. Nel diciassettesimo giorno le cellule mesodermiche hanno occupato tutto lo spazio disponibile tra endoderma ed ectoderma, tranne che in corrispondenza delle membrane faringea e cloacale, dove è del tutto assente. Le cellule che penetrano attraverso il nodo di Hensen si muovono in linea retta verso la membrana faringea creando il cosiddetto processo cefalico all'interno del foglietto mesodermico, e una depressione dal lato ectodermico chiamata fossetta primitiva. Questa fossetta si prolunga all'interno del processo cefalico trasformandolo in un tubo cavo (canale cordale), spingendosi fino al sacco vitellino, e permettendo la comunicazione tra questi e la cavità amniotica, attraverso il canale neurenterico o di Lieberkühn.
Durante la formazione della notocorda le cellule immediatamente sovrastanti diventano più alte e voluminose creando una regione nell'ectoderma più spessa, la placca neurale. Questa inizialmente ha una forma ovoidale che si estende a mano a mano che il processo cefalico avanza. Al diciannovesimo giorno, quando la notocorda si è formata completamente, i bordi della placca neurale si innalzano formando le pieghe neurali che si incurvano sempre più e, al ventunesimo giorno, convergono e si chiudono dando origine al tubo neurale e alla cresta neurale, che è la parte della doccia che non si chiude nel tubo. Al 24º giorno il tubo neurale è chiuso lungo l'asse e anche le estremità (neuropori) sono chiuse, quella posteriore in leggero ritardo. La cresta neurale, considerata un quarto foglietto, migra in diverse direzioni, formando le più disparate categorie di cellule come neuroni, neuroglia, melanociti e parte del derma, cartilagini della testa, muscoli dell'iride, la pars midollare del surrene e il setto del cuore.
Nel diciottesimo giorno, le cellule del mesoderma più vicine al processo cefalico proliferano attivamente dando origine a ispessimenti paralleli, il mesoderma parassiale. In principio si formano 42 coppie di somitomeri, di cui le prime 7 si uniscono tra loro, le ultime 5 degenerano, mentre le restanti 32 si frammentano in blocchi pieni chiamati somiti. Questi creano una cavità centrale, il miocele. Le cellule rivolte verso il tubo neurale costituiscono lo sclerotomo, che vanno a formare vertebre, coste e scapole; quelle esterne formano il dermomiotomo, che si suddivide in dermatomo, che è parte del derma, e miotomo, che darà origine ai muscoli del tronco e degli arti.
Nel mesoderma laterale, invece, si creano delle cavità che confluiscono separandolo in somatopleura, a contatto con l'ectoderma, e splancnopleura, che ricopre l'endoderma. Il tratto di mesoderma che collega il parassiale al laterale si ispessisce a sua volta in due cordoni paralleli, il mesoderma intermedio. Il celoma, dopo l'avvolgimento dell'amnios, viene racchiuso in cavità: quella pleurica, pericardica e peritoneale.
Sviluppo del Cuore e dei Vasi Sanguigni
Il cuore e i vasi sanguigni derivano dagli angioblasti, cellule mesenchimali che derivano dalla splancnopleura. Gli angioblasti si aggregano, invece, in vescicole sempre più grandi formando i cordoni angioblastici che accrescendosi e unendosi tra loro creano vasi sanguigni distribuiti uniformemente. La prima rete di vasi si forma nel sacco vitellino, poi nel peduncolo, poi nel corion e finalmente si uniscono agli abbozzi del cuore. Nell'area cardiogena le cellule del mesenchima formano due cordoni endocardici, destro e sinistro, che creano le cavità al loro interno e si collegano tra loro a seguito dei ripiegamenti della quarta settimana in un tubo solo, l'endocardio. Successivamente altre cellule, prevalentemente contrattili, si associano al tubo cardiaco generando il miocardio, separato dall'endocardio dalla gelatina cardiaca. Un terzo gruppo di cellule ricopre il miocardio, formando l'epicardio.
I Piegamenti dell'Embrione e la Definizione della Forma
Nella quarta settimana avviene la trasformazione dell'embrione da una struttura essenzialmente piatta ad una cilindrica, e la crescita della cavità amniotica, e quindi dell'ectoderma, più rapida rispetto al resto dell'embrione, e comunque in senso longitudinale. Tale espansione si traduce in uno "strabordo" dell'ectoderma sul resto del corpo, creando così una piega cefalica e una piega caudale, che gli abbozzi anteriori si portano sotto l'endoderma e ruotano su loro stessi; la rotazione a metà della quarta settimana è di 180°. Le pieghe si sviluppano anche in senso trasversale, con l'effetto di deporre uno strato di amnios attorno all'intero embrione e di ripiegare appunto i foglietti ruotando gli abbozzi. Le due pieghe danno origine quindi all'intestino primitivo, il cui lume è formato dal sacco vitellino rimasto nella piega dell'endoderma. La restante parte dell'endoderma crea il dotto vitellino.
Come nasce un essere umano: l'incredibile percorso dalla fecondazione al parto
L'Embodiment dello Spazio: Una Prospettiva Olistica e Cosciente
Al di là della pura descrizione anatomica e cronologica, esiste una prospettiva che vede lo sviluppo embrionale non solo come l'embodiment della struttura, ma anche come l'embodiment dello spazio. In questo senso, il "luogo" primordiale di cui si parla è il vuoto, il nostro primo embodiment, prima ancora di dare corpo alla struttura, è quello che noi facciamo in relazione allo spazio vuoto. Durante la nostra vicenda embriologica, noi passiamo attraverso una serie di spazi. Bonnie B. suggerisce che quando si entra in uno stato meditativo o si è partecipi di forme di sottile movimento interiore, come nelle pratiche del Body-Mind Centering® - o in altre come il Continuum, il Movimento Autentico, la danza Sufi, la Contact Improvisation, il Katsugen Undo - s’arriva a un punto in cui ci si connette a un senso primordiale dello spazio: leggerezza, agio, flusso.
Il Sacco Vitellino, la Cavità Peritoneale e la Borsa Omentale
Bonnie B. nota che, tornando al percorso evolutivo del feto, alla fine della terza settimana, il tratto digerente, il cervello e il cuore hanno cominciato a svilupparsi. Alla fine della quarta settimana, "ingurgitato" il sacco vitellino nella sua propria pancia, l’embrione comincia a curvarsi prendendo una forma a C. Il sacco vitellino, una volta esaurita la maggior parte della sostanza nutritiva, diventa, così si sente, il sacco peritoneale, che, nell’adulto già completamente sviluppato, è sede della maggior parte degli organi addominali. Questo sacco peritoneale è unodei punti su cui concentrarsi nell'embodiment dello spazio a livello embriologico. Da adulti, la membrana peritoneale della cavità addominale, che si forma dal sacco vitellino, contiene gli organi interni e il liquido sieroso. Se rafforziamo queste membrane e se cominciamo a sentire lo spazio che accoglie gli organi, sentiamo di esserci realmente, liberi, senza cose da fare.
Una particolare cavità descritta da Bonnie B. è la borsa omentale, uno spazio formatosi, durante la vita fetale, attraverso la rotazione del tratto digerente, situata dietro lo stomaco, davanti al pancreas e sotto il fegato. "Borsa" significa spazio. Nella parte posteriore si trova il forame epiploico, o di Winslow, una cavità che connette la borsa omentale al grande sacco del peritoneo, dove si trovano quasi tutti gli organi interni. Questo forame omentale è chiamato anche forame premistoppa, e prende il nome dall'anatomico Jacob B. Winslow. L'epiplon deriva dal greco "epi" (sopra) e "ploon" (che galleggia), nel senso di '(membrana) fluttuante al di sopra'. È una ripiegatura del peritoneo, derivata dal mesogastrio e dal mesocolon, con funzione di protezione della massa delle anse dell'intestino tenue, che congiunge tra loro i visceri dell'addome; tesa fra i visceri, prende nome di “epiploon (o legamento) gastro-epatico“, detto anche “piccolo omento“, e “epiploon (o legamento) gastro-colico“, detto anche grande omento.
Respirazione, Energia e Longevità: La Cavità Midollare
L'esplorazione di questi spazi interni può essere intrapresa anche attraverso pratiche come la respirazione. Bonnie B. condivide la sua esperienza nel respirare nella borsa omentale, notando che con uno sforzo minimo, il forame epiploico si apre e il respiro passa in un'altra cavità, e pian piano attraverso varie cavità. L'obiettivo è quello di respirare "sfericamente", riempiendo tutta la zona: davanti, di dietro, lateralmente, sopra e sotto. Questo tipo di respiro diventa un mare di chi, muovendosi a onda. Bonnie B. ha scoperto che le membrane diventano così forti che il movimento non è più muscolare, ma è una tenuta dello spazio, della cavità peritoneale che contiene gli organi. È sorprendente come l'utilizzo della membrana, che lei sente essere ciò che rimane del sacco vitellino, sia ancora presente e funzionale.
Questo concetto si estende anche alla cavità midollare. Bonnie B. suggerisce che quando si sviluppa energia nel tantien, essa raggiunge la coda, va su per la spina dorsale, scende fino ai piedi, e passa attraverso il corpo intero, raggiungendo il midollo osseo. Quando si trova nel midollo, forma un rivestimento, una sorta di fodera, nello spazio interno dell’osso, nella cavità midollare. Quando questo accade, le porte della buona salute e della longevità si aprono. La scienza contemporanea, nel suo studio dello sviluppo umano, definisce le prime fasi dell'organismo umano come zigote, morula, blastula e gastrula. L'Organizzazione mondiale della sanità ha stabilito che dopo la fecondazione e fino all'incirca al quattordicesimo giorno, il prodotto del concepimento potrebbe dividersi dando vita a un parto con più nascituri. Questa e altre considerazioni portano alcune correnti di pensiero ad affermare che non si può parlare di embrione nel senso di futura persona prima del 14º giorno di gestazione. Tuttavia, tutte le fasi dello sviluppo sono regolate, dall'interno, dai geni, ereditati dai genitori e, dall'esterno, dalla complessa interazione biochimica con la mucosa uterina e con l'organismo della madre. Possono incidere sui fattori esterni l'alimentazione della madre ed eventuali farmaci da lei assunti durante la gravidanza.