La comprensione dei meccanismi con cui le cellule rispondono agli stimoli esterni è fondamentale per svelare le complessità della biologia e della farmacologia. Tra questi meccanismi, la desensibilizzazione dei recettori occupa un posto di rilievo, rappresentando una proprietà intrinseca di molti sistemi di segnalazione cellulare. Questo fenomeno fisiologico, che porta a una ridotta capacità di risposta a uno stimolo persistente, si manifesta in diverse forme, le più discusse delle quali sono la desensibilizzazione omologa ed eterologa. Sebbene entrambe descrivano una perdita di risposta recettoriale, differiscono significativamente nei loro meccanismi d'azione e nelle conseguenze. Parallelamente, in un contesto completamente diverso ma ugualmente cruciale per la vita umana, i concetti di "omologo" ed "eterologo" trovano applicazione nella Procreazione Medicalmente Assistita (PMA), descrivendo la provenienza dei gameti utilizzati per creare una nuova vita. Analizzare queste due sfere, quella molecolare e quella riproduttiva, offre una prospettiva unica sulle sfumature del linguaggio scientifico e biologico.
La Desensibilizzazione: Un Meccanismo di Regolazione Cellulare
La desensibilizzazione è un processo biologico attraverso il quale una cellula o un organismo riduce la propria risposta a uno stimolo, come un farmaco, un ormone o un neurotrasmettitore, dopo un'esposizione prolungata o ripetuta. Questo meccanismo è vitale per prevenire l'iperstimolazione e mantenere l'omeostasi cellulare.
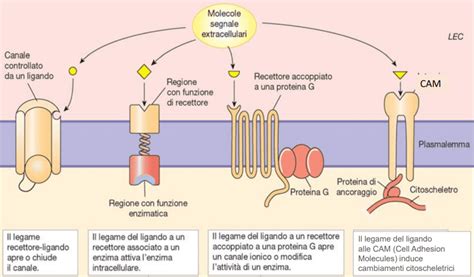
Nel caso dei recettori-canale, la desensibilizzazione è una proprietà intrinseca che determina la riduzione della capacità del recettore di andare incontro a quelle modificazioni conformazionali necessarie al funzionamento del suo canale intrinseco. È un fenomeno causato dall’attivazione continua del recettore-canale ed è completamente reversibile appena il farmaco (o il ligando in generale) si stacca dal recettore-canale. In sintesi, quando un ligando non si stacca dal recettore in tempi ottimali determina desensitizzazione: il ligando è legato al recettore ma il canale è chiuso.
Normalmente quando un recettore-canale viene attivato da un agonista, il canale passa da uno stato di riposo ad uno stato attivo: il canale si apre e conduce gli ioni. Il canale passa spontaneamente allo stato inattivo, ma se il farmaco agonista persiste, il canale permane per un tempo più lungo in uno stato conformazionale che corrisponde ad uno stato inattivo del canale che è lo stato che non conduce. Il passaggio del canale dallo stato attivo allo stato inattivo-desensitizzato è modulato da protein-chinasi (di tipo A o di tipo C) che possono fosforilare la parte intracellulare del recettore. L’esempio caratteristico è quello dei terminali nervosi della giunzione neuromuscolare che, quando stimolati ad alta frequenza, non solo liberano il neurotrasmettitore ma anche un peptide (CGRP - peptide correlato al gene della calcitonina-) che attiva dei meccanismi che portano ad un aumento della concentrazione intracellulare di cAMP che attiva la PKA che fosforila la parte interna del recettore favorendo lo stato desensitizzato di questo canale. Tale peptide ha un ruolo importante nel danno che si instaura dopo stimolazione cronica con questi farmaci.
Si potrebbe pensare che il passaggio del canale dallo stato attivo allo stato inattivo-desensitizzato sia accompagnato da una riduzione di affinità del neurotrasmettitore o del farmaco nei confronti del recettore. In realtà avviene l’esatto contrario. Un canale nello stato desensitizzato è molto più affine al legame con il ligando agonista. L’acetilcolina ha una attività per lo stato di riposo del canale di circa 100 µM, cioè necessita di tale quantità per legare il 50% dei recettori nicotinici, mentre nello stato inattivato l’attività dell’acetilcolina passa a 5 µM, che allo stato desensitizzato passa ad 1µM.
La desensibilizzazione è un fenomeno importante perché ha due implicazioni farmacologiche: è un fenomeno che si instaura rapidamente e reverte altrettanto rapidamente e completamente; la rapidità consente l’uso di alcuni farmaci come la succinilcolina che è utilizzata per bloccare la trasmissione neuromuscolare (non è un anestetico ma è usato con l’antestetico perché l’anestesia si potrebbe estinguere oltre un certo grado; per evitare ciò si associano questi farmaci che bloccano la trasmissione neuromuscolare con desensitizzazione). La succinilcolina, come la tubocurarina, fa parte dei curari che chimicamente sono alcaloidi naturali; la tubocuranina, però, è un bloccante neuromuscolare che non agisce per desensitizzazione ma agisce con meccanismo competitivo nei confronti dell’acetilcolina. Il farmaco lega il recettore nicotinico in un certo sito in cui si lega l’acetilcolina e per competizione blocca la trasmissione neuromuscolare. La succinilcolina, invece, opera per desensitizzazione attraverso un meccanismo depolarizzante, infatti blocca la trasmissione neuromuscolare ma è un farmaco agonista del recettore nicotinico: lo attiva ma a differenza dell’ACh non è degradata dall’ACh-esterasi e la sua persistenza sul recettore nicotinico rende il canale desensitizzato. In un paziente in cui viene somministrata succinilcolina vi sarà una prima fase di attivazione e dopo si ha la paralisi della muscolatura per blocco.
Desensibilizzazione Omologa: Specificità d'Azione
La desensibilizzazione omologa è definita come la perdita di attività dell’agonista che è specifica per il recettore che viene attivato. Questo significa che, quando un recettore viene stimolato in modo continuativo, la sua capacità di rispondere si riduce, ma questa riduzione non influenza altri tipi di recettori presenti sulla stessa cellula, a meno che non condividano esattamente lo stesso destino molecolare.
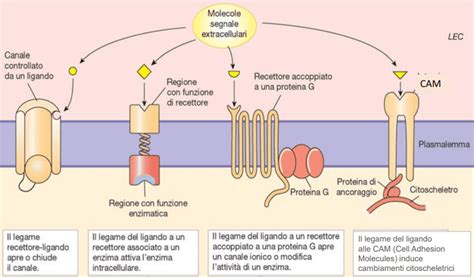
Nel caso dei recettori-canale, come accennato, la desensibilizzazione omologa avviene quando il ligando persiste nel legame, mantenendo il canale in uno stato inattivo. Per i recettori accoppiati a proteine G, la desensibilizzazione omologa si verifica quando l'attivazione prolungata di un recettore porta alla sua inattivazione specifica. Questo meccanismo è spesso mediato da modificazioni post-traduzionali del recettore stesso. La fosforilazione del recettore su residui di serina e treonina è una tappa chiave, spesso mediata da protein-chinasi specifiche per i recettori accoppiati alla proteina G (GRK). Questa fosforilazione aumenta l'affinità del recettore per una proteina citoplasmatica chiamata beta-arrestina, che, traslocando sulla membrana, determina il distacco del recettore dalla proteina G. A questo punto, il recettore può essere internalizzato in endosomi dopo essere stato rivestito da clatrina. Il processo di fosforilazione, che è la prima tappa del processo, è mediato da protein-kinasi A e C a livello della terza ansa citoplasmatica; è seguito dalla perdita della capacità del recettore di attivare la proteina G (disaccoppiamento tra recettore e proteina G); non viene così trasdotto il segnale fino ad arrivare alla riduzione del numero di recettori di membrana col fenomeno seguente della down-regulation cioè quel fenomeno che porta alla riduzione del numero dei recettori di membrana.
Desensibilizzazione Eterologa: Interconnessioni Segnaletiche
A differenza della desensibilizzazione omologa, la desensibilizzazione eterologa si estende anche ad altri recettori che utilizzano la stessa via di trasduzione del segnale o gli stessi effettori. Ciò implica che l'attivazione prolungata di un sistema recettoriale può indurre la desensibilizzazione di un altro sistema recettoriale presente sulla stessa membrana cellulare.
Questo fenomeno è particolarmente rilevante quando diversi tipi di recettori convergono su effettori comuni, come le proteine G o le chinasi a valle nella cascata di segnalazione. Ad esempio, se un farmaco agonista attiva un recettore accoppiato a proteine G che, a sua volta, attiva la protein-chinasi A (PKA), e sulla stessa cellula è presente un altro tipo di recettore che, una volta attivato, porta anch'esso all'attivazione della PKA, allora la desensibilizzazione del primo recettore potrebbe influenzare la risposta del secondo, anche se quest'ultimo non è stato direttamente stimolato.
Un esempio di desensibilizzazione eterologa si osserva nel trattamento cronico con farmaci attivi sulle proteine G, che può indurre un fenomeno di tolleranza farmacologica su base farmacodinamica. Questa tolleranza è prettamente legata al meccanismo d’azione dei farmaci e correlata alle modificazioni dei recettori che vengono esposti cronicamente ad un farmaco. È un fenomeno che reverte con la sospensione della terapia. Le conseguenze di tolleranza in corso di terapia sono molteplici: la stessa dose di farmaco può non essere sufficiente a controllare la malattia e occorre aumentare il dosaggio per ottenere lo stesso effetto; ovviamente si potrà andare incontro ad un nuovo stato di tolleranza. Inoltre, vi è la possibilità che la sospensione rapida della malattia si accompagni ad un effetto rimbalzo dovuto ad uno squilibrio recettoriale. Ad esempio, se il soggetto è trattato con broncodilatatori, agonisti del recettore b-adrenergico, e si sospende bruscamente la terapia, il soggetto può andare incontro a crisi asmatica; se il soggetto è trattato con farmaci antiipertensivi che attivano un recettore adrenergico, la sospensione brusca della terapia può causare una crisi ipertensiva (come nel caso di utilizzo di clonidina). Alla base di questi fenomeni c’è la desensitizzazione del recettore, cioè la perdita della capacità di trasduzione del segnale.
Meccanismi Molecolari Condivisi e Distinti
I meccanismi responsabili della desensibilizzazione, sia omologa che eterologa, includono la fosforilazione del recettore su residui di serina e treonina e l’internalizzazione del complesso recettore-ligando. Dopo che il complesso recettore-ligando è stato fosforilato ed internalizzato, può essere degradato o riciclato sulla membrana (risensitizzazione). Il dimero beta/gamma recluta chinasi specifiche dei recettori accoppiati alla proteina G (GRK); le GRK fosforilano il recettore su residui di serina e treonina sulla parte intracellulare del recettore. Questa fosforilazione fa aumentare l’affinità del recettore per una proteina citoplasmatica denominate beta-arrestina che trasloca sulla membrana e determina il distacco del recettore dalla proteina G. A questo punto il recettore può essere internalizzato in endosomi dopo essere stato rivestito da clatrina. Il recettore può essere degradato da enzimi lisosomiali (quindi si ha down-regulation) o può ritornare sulla membrana cellulare (risensitizzazione).
Nel caso dei recettori-canale, la desensibilizzazione è una proprietà intrinseca che determina la riduzione della capacità del recettore di andare incontro a quelle modificazioni conformazionali necessarie al funzionamento del suo canale intrinseco. Questo avviene per attivazione continua del recettore-canale. La desensibilizzazione può essere rapida (componente I, con il 50% dei recettori che si inattiva in poche centinaia di millisecondi) o lenta (componente D, dove il 50% dei recettori si inattiva in molto più tempo).
Esempi di molecole che inducono desensibilizzazione includono la nicotina. Un altro esempio è dato dalla nicotina, un alcaloide derivato dalla Nicotiana Tabacum, che ad alte dosi è un antagonista del recettore colinergico e la sua funzione cronica porta alla desensitizzazione dei recettori nicotinici neuronali. Questo meccanismo determina una riduzione persistente della trasmissione che causa un fenomeno paradosso (nell’assunzione cronica di nicotina) che è la up-regulation. Gli stessi recettori nicotinici desensitizzati per l’uso cronico di nicotina vanno incontro ad una maggiore sintesi per cui ne aumenta il numero a livello del vallo sinaptico colinergico.
Intossicazione acuta da anticolinesterasici: Questi sono farmaci che inibiscono l’ACh-esterasi, sono irreversibili e hanno interesse terapeutico ma soprattutto tossicologico. Questi inibitori sono composti che appartengono alla classe degli organofosforici (insetticidi e gas nervini) che legano in maniera irreversibile l’ACh-esterasi e di conseguenza inducono una paralisi muscolare; la loro pericolosità è aggravata dalla liposolubilità: possono così attraversare la barriera emato-encefalica con effetti sul SNC. La paralisi è preceduta da fascicolazioni muscolari come per la succinilcolina, perché il blocco dell’ACh-esterasi mantiene proprio l’ACh nel vallo sinaptico e si hanno quei segni da iperattivazione del recettore nicotinico seguiti da paralisi.
Procreazione Medicalmente Assistita: Omologo ed Eterologo
Nel campo della riproduzione umana, i termini "omologo" ed "eterologo" assumono un significato completamente diverso, riferendosi alla provenienza genetica dei gameti utilizzati nei percorsi di Procreazione Medicalmente Assistita (PMA).
La Procreazione Medicalmente Assistita (PMA) consiste in una serie di procedure e tecniche che comportano il trattamento di ovociti umani, di spermatozoi o embrioni allo scopo di realizzare una gravidanza. Le tecniche di PMA sono indicate per le coppie con infertilità comprovata - in cui un partner o entrambi presentano problematiche accertate da un iter medico - o in caso di infertilità idiopatica. La procreazione assistita entra in gioco quando la coppia desidera avere un figlio ma non riesce a ottenere una gravidanza tramite la fecondazione naturale.

Fecondazione Omologa: L'Unione dei Gameti della Coppia
La fecondazione omologa è una tecnica di PMA che prevede lo sviluppo e l’eventuale impianto in utero di un embrione ottenuto con l’impiego di gameti (spermatozoi e ovuli) appartenenti alla coppia richiedente. In questo caso, il patrimonio genetico del nascituro deriverà da entrambi i genitori biologici.
Generalmente si ricorre alla fecondazione omologa in tutti quei casi in cui c’è un’infertilità maschile di grado lieve o moderato, disfunzioni sessuali, ripetuti tentativi di concepimento per più di due anni non andati a buon fine, in caso di disfunzioni ovulatorie o di infertilità idiopatica. È il primo step da provare, prima di ricorrere alla donazione, laddove non vi sia una chiara definizione della causa del mancato concepimento.
Le tecniche di PMA sono suddivise in I, II e III livello a seconda della loro complessità e della necessità o meno di anestesia.Tecniche di primo livello: inseminazione intrauterina (IUI). La IUI è la tecnica di PMA meno invasiva, svolta in regime ambulatoriale, nel giro di poche ore. Prevede l’iniezione nella cavità uterina di liquido seminale durante i picchi di ovulazione, monitorati tramite ecografia transvaginale. Quando l’ovulazione non avviene spontaneamente, viene stimolata farmacologicamente (stimolazione ovarica) sotto stretta osservazione del medico. Nell'inseminazione intrauterina nella fecondazione omologa, il liquido seminale appartiene al partner e viene prelevato il medesimo giorno dell’inseminazione. Lo sperma è poi preparato in laboratorio e iniettato nella cavità uterina, lasciando fare alla natura il proprio corso.
Tecniche di secondo livello: fecondazione in vitro (FIVET) o microiniezione intracitoplastica (ICSI). Queste tecniche sono più invasive per la donna. Si basano sul prelievo di uno o più ovociti dall’utero, che verranno poi fecondati in vitro. Nelle tecniche di secondo livello per la fecondazione omologa, la cellula uovo appartiene alla donna della coppia e non a una donatrice. Tutto l’iter diagnostico, medico e burocratico non coinvolgerà terze parti e sarà valutato sulle problematiche di fertilità riscontrate in fase di indagine.
Tecniche di terzo livello: prelievo chirurgico degli spermatozoi (MESA, TESA, TESE). In questa categoria rientrano le tecniche che prendono il nome di Mesa, Tesa e Tese e vengono messe in campo in caso di grave infertilità e azoospermia, dovuta a ostruzione delle vie seminali o a un mancato processo di maturazione degli spermatozoi. Le tecniche di terzo livello sono valide solo in caso di fecondazione omologa e in tutte si preleva il liquido seminale in modo invasivo. L’asportazione è seguita dalla fecondazione in vitro e dal successivo impianto dell’embrione nella cavità uterina della madre biologica.
Fecondazione Eterologa: Il Ricorso alla Donazione
Diverso è il punto di partenza per chi decide di orientarsi verso la fecondazione eterologa. Qui l’infertilità è assoluta per mancanza di gameti in uno dei due partner o in entrambi. Sebbene entrambi i componenti della coppia per la legge siano a tutti gli effetti genitori, almeno uno dei due gameti non appartiene alla coppia. Quindi, il patrimonio genetico sarà di un solo genitore.
In Italia, il ricorso a gameti esterni è permesso laddove non vi siano altre terapie efficaci in grado di risolvere le cause di infertilità o sterilità. L’eterologa nel nostro Paese è rimasta vietata fino al 2014, quando una sentenza della Corte Costituzionale ha di fatto modificato la Legge 40 a favore di chi, impossibilitato ad usare i propri gameti, ha potuto fare ricorso a materiale biologico proveniente da donatore.
Se l’infertilità riguarda la donna si parla di ovodonazione, mentre se è dell’uomo, di eterologa maschile. Possono accedere all’eterologa le donne che hanno disturbi ormonali e non rispondono alla stimolazione ormonale, oppure quelle in età avanzata (ma non troppo) che hanno una ridotta o assente riserva ovarica o se portatrici di un difetto genetico. Le situazioni in cui è possibile ricorrere a questa specifica metodica sono molto selezionate: si tratta soprattutto di donne affette da gravi patologie ginecologiche, come lesioni tumorali che necessitano interventi chirurgici o chemio/radioterapici.
Nell’inseminazione intrauterina nella fecondazione eterologa, grazie alla legge 40 del 2004, la coppia può affidarsi a un centro di fecondazione assistita per ottenere il liquido seminale di un donatore, che sarà selezionato da équipe specializzate, tenendo conto di etnia, gruppo sanguigno e caratteristiche genetiche del donatore.
Per quanto riguarda le tecniche di secondo livello (FIVET e ICSI) nella fecondazione eterologa, la procedura medica è la medesima, ma l’ovulo appartiene a una donatrice, mentre il liquido seminale proviene dal papà biologico o viceversa.
Normativa e Aspetti Sociali in Italia
La Procreazione Medicalmente Assistita è regolata dalla Legge n° 40 del 2004 e successive modifiche che stabiliscono sostanzialmente chi può accedervi ed in quali casi. In via generale la normativa fa riferimento a “coppie di maggiorenni di sesso diverso, coniugate o conviventi, in età potenzialmente fertile, entrambi viventi“.
In Italia, in nessun caso la coppia che riceve ovociti o spermatozoi può conoscere l’identità del donatore, né il donatore quella dell’eventuale bimbo. I “dati clinici del donatore/donatrice potranno essere resi noti al personale sanitario solo in casi straordinari, dietro specifica richiesta e con procedure istituzionalizzate, per eventuali problemi medici della prole, ma in nessun caso alla coppia ricevente. Amici, familiari e parenti non sono candidabili a donatori. Inoltre, è “previsto per i donatori e per le donatrici un limite massimo di 10 nati. La donatrice dovrà avere tra i 20 e i 35 anni e il donatore tra i 18 e i 40.
Purtroppo in Italia, per vari motivi i donatori di gameti sono ancora pochi. Questo vale soprattutto per gli ovuli femminili, considerato che per ottenerli con un prelievo c’è un percorso lungo di stimolazione ovarica a cui sottoporsi che può essere pesante. È possibile dunque far ricorso a criobanche all’estero. Questo però non significa fare “viaggi della speranza”. Basta rivolgersi ad un centro per la fertilità qualificato e di lunga esperienza vicino casa. Saranno i professionisti che vi lavorano a contattare una criobanca per reperire materiale biologico compatibile con la coppia, da impiegare nella fecondazione assistita.
Conclusioni Differenziate: Dai Meccanismi Cellulari alla Vita
La distinzione tra desensibilizzazione omologa ed eterologa risiede nella specificità dell'effetto: la prima colpisce il recettore direttamente attivato, mentre la seconda può influenzare vie di segnalazione condivise da recettori diversi. Questa fine regolazione cellulare è essenziale per la sopravvivenza e l'adattamento dell'organismo.
Parallelamente, nel contesto della riproduzione assistita, i termini omologo ed eterologo definiscono la provenienza dei gameti, un aspetto cruciale che determina la composizione genetica del nascituro e le implicazioni legali e sociali della genitorialità. In entrambi i campi, sebbene apparentemente distanti, la terminologia sottolinea la complessità dei processi biologici e la necessità di una comprensione dettagliata dei meccanismi sottostanti. La scelta tra fecondazione omologa o eterologa, come la comprensione della desensibilizzazione recettoriale, richiede un'analisi approfondita dei fattori individuali e delle specifiche circostanze. La scienza continua a evolversi, offrendo nuove prospettive e soluzioni in entrambi questi ambiti vitali.
tags: #desensibilizzazione #omologa #ed #eterologa