L'emopoiesi, nota anche come ematopoiesi, è il processo biologico fondamentale che governa la formazione e la maturazione di ogni singolo tipo di cellula sanguigna. Queste cellule, essenziali per la vita, originano da precursori specifici e attraversano un complesso percorso di differenziazione. Il termine stesso, derivante dal greco aima (sangue) e poieo (creare), ne sottolinea la funzione vitale. All'interno di questo vasto processo, la mielopoiesi rappresenta una delle direzioni principali di maturazione delle cellule staminali emopoietiche, portando alla genesi dei globuli bianchi (granulociti e monociti), dei megacariociti (responsabili della produzione di piastrine) e dei globuli rossi. Questo processo non è statico, ma si evolve dinamicamente, spostandosi attraverso diverse sedi anatomiche durante le fasi cruciali dello sviluppo fetale, adattandosi alle mutevoli esigenze dell'organismo in crescita.
L'Alba dell'Emopoiesi: Il Sacco Vitellino e la Fase Vitellina
Il viaggio della formazione delle cellule del sangue inizia in modo sorprendentemente precoce nello sviluppo embrionale. L’emopoiesi, inclusa la prima, rudimentale forma di mielopoiesi, ha inizio circa 21 giorni dalla fecondazione. Questo momento coincide con circa 2 settimane dall’annidamento della blastocisti, una fase embrionale propria dei mammiferi, all’interno della mucosa uterina. Contrariamente a quanto si potrebbe pensare, le prime fasi di questo processo vitale non avvengono direttamente nell'embrione, ma al di fuori di esso. La localizzazione iniziale è nel mesenchima extraembrionale, specificamente nella parete del sacco vitellino primitivo. Questa particolare zona è il terreno fertile dove, nello spessore del mesoderma, si formano i primi isolotti emopoietici di Wolff.
In questi isolotti, si assiste alla nascita delle prime cellule eritroidi. È importante notare che queste cellule sono ancora dotate di nucleo, una caratteristica che le distingue nettamente dai globuli rossi maturi che osserveremo successivamente, i quali sono anucleati per massimizzare la loro funzione di trasporto di gas. Circa 4 settimane dopo la fecondazione, un momento significativo nello sviluppo embrionale è segnato dalla formazione di un cuore pulsante. Questa pietra miliare permette a queste prime cellule eritroidi nucleate di iniziare a circolare attivamente entro una rete vasale primitiva che si sta abbozzando. La fase vitellina dell’emopoiesi è cruciale per la sopravvivenza precoce dell'embrione, fornendo l'ossigeno e i nutrienti necessari per il suo rapido sviluppo. Tuttavia, la sua importanza diminuisce progressivamente. L’emopoiesi vitellina inizia a ridursi verso la 7a settimana di vita embrionale, sebbene continui a un ritmo più contenuto fino alla 12a settimana. Durante questo periodo, si verifica una transizione e una sovrapposizione con la fase successiva dell'emopoiesi, quella epatica, preparandosi a lasciare il posto a organi più specializzati per questa funzione. Sebbene la produzione di globuli rossi sia predominante in questa fase, i progenitori emopoietici presenti nel sacco vitellino hanno la capacità di dare origine anche a cellule mieloidi primitive, fondamentali per le prime difese innate dell'embrione.
Il Fegato Fetale: Un Centro Emopoietico Transitorio
Con l'avanzare dello sviluppo, l'onere della produzione di cellule sanguigne si sposta verso un nuovo organo: il fegato fetale. L’emopoiesi epatica, che include la mielopoiesi fetale, ha inizio verso la fine dell’8a settimana di sviluppo embrionale, subentrando gradualmente alla funzione del sacco vitellino. In questa fase, il fegato non è solo l'organo principale per il metabolismo e la detossificazione, ma assume un ruolo preponderante come sede di emopoiesi. Il suo ambiente ricco di fattori di crescita e la sua estesa vascolarizzazione lo rendono ideale per supportare la proliferazione e la differenziazione delle cellule staminali emopoietiche.
Durante la fase epatica, il fegato fetale produce una gamma più ampia di cellule ematiche rispetto al sacco vitellino. Non si limita solo agli eritrociti, ma è anche un sito attivo per la produzione di leucociti, inclusi i progenitori mieloidi, e trombociti. Questo è un periodo di intensa proliferazione cellulare, essenziale per la crescita esponenziale del feto. Tuttavia, la supremazia del fegato come organo emopoietico è transitoria. La sua attività diminuisce rapidamente verso il 5° mese di gestazione. Questa diminuzione è strategicamente coordinata con l'emergere di un'altra sede emopoietica, che diventerà la sede definitiva della produzione di sangue. L’emopoiesi epatica continua a un ritmo ridotto fino alla 1a settimana di vita extrauterina, sovrapponendosi a quella che sarà l’emopoiesi definitiva. Questo periodo di transizione e sovrapposizione è un esempio eloquente della flessibilità e della ridondanza del sistema emopoietico durante lo sviluppo, garantendo che non vi siano interruzioni nella fornitura di cellule sanguigne vitali. La mielopoiesi in questa fase è cruciale per la costituzione di un sistema immunitario fetale, sebbene ancora immaturo.
SISTEMA EMOLINFOPOIETICO - PARTE SECONDA -
Il Midollo Osseo: La Sede Definitiva della Mielopoiesi
Il midollo osseo emerge come la sede primaria e definitiva dell'emopoiesi, inclusa la mielopoiesi, a partire dalla seconda metà della gestazione. L’emopoiesi definitiva inizia verso il 5° mese di vita fetale e, una volta stabilita, continua per tutta la vita postnatale. In questo momento, il midollo osseo del feto diventa emopoietico, ovvero capace di emopoiesi, e questa fase si sovrappone a quella epatica fino a prenderne il pieno controllo. La sua localizzazione è caratteristicamente all'interno delle cavità ossee, in particolare nel tessuto molle e spugnoso. Nell'essere umano adulto normale, le cellule ematiche sono formate in modo continuo e regolare in questa sede specializzata.
Inizialmente, durante il periodo fetale e nell'infanzia, gran parte dello scheletro contiene midollo osseo emopoieticamente attivo. Tuttavia, gli spazi occupati da midollo emopoietico si riducono progressivamente dall'infanzia all'età adulta, fino ad essere confinati prevalentemente alla parte centrale dello scheletro. Alla pubertà, l’emopoiesi si verifica principalmente nello sterno, nelle vertebre, nelle ossa iliache (del bacino) e nelle costole. Questo cambiamento nella distribuzione del midollo attivo riflette un'ottimizzazione dell'organismo, concentrando la produzione di sangue in siti strategicamente protetti e funzionalmente efficienti. Il midollo osseo è il principale produttore di tali elementi, nello specifico quello localizzato all’interno dello sterno, delle vertebre, delle costole e delle scapole. Il sistema emopoietico permette di aggiornare continuamente i globuli rossi, le piastrine e i globuli bianchi (granulociti, monociti e linfociti) inclusi nel sangue. La maggior parte delle cellule ematiche è incapace di ulteriore divisione (fase G0) e, avendo vita relativamente breve, è rimpiazzata continuamente da nuovi elementi provenienti dal midollo osseo. Questo incessante ricambio sottolinea l'importanza del midollo osseo come fabbrica di cellule sanguigne perenne e adattabile. All'interno del midollo osseo, le cellule staminali pluripotenti danno origine ai precursori dei diversi tipi di cellule ematiche, garantendo una mielopoiesi robusta e continua.
La Cellula Staminale Emopoietica Pluripotente: Il Cuore della Mielopoiesi
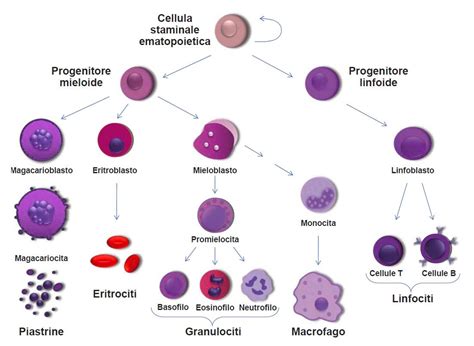
Al centro di tutto il processo emopoietico, e quindi mielopoietico, si trova una cellula straordinaria: la cellula staminale emopoietica pluripotente. L’emopoiesi ha origine proprio in queste cellule staminali, cellule primitive classificate sulla base della loro potenzialità di differenziarsi in varie linee cellulari. Globuli rossi, globuli bianchi e piastrine, tutti elementi corpuscolati del sangue, derivano da un'unica cellula staminale emopoietica pluripotente. Questa è la prima di una serie di tappe, sequenziali e ordinate, di crescita e maturazione cellulare. La differenziazione cellulare, infatti, è quel particolare processo di maturazione e specializzazione delle cellule dell’organismo che, dopo le prime divisioni cellulari durante la fase embrionale, diventano sempre più "competenti" nel compiere una determinata funzione, assumendo quindi caratteristiche (ad esempio, la forma cellulare e nucleare) che le rendono distinguibili le une dalle altre.
Le attuali conoscenze si basano su due nozioni fondamentali: la prima è che le cellule del sangue (globuli rossi, globuli bianchi e piastrine) hanno tutte origine da un unico tipo di cellula staminale emopoietica pluripotente; la seconda è che la maturazione di quest'ultima può procedere lungo due direzioni principali: la linea linfoide e la linea mieloide, attivando rispettivamente la linfopoiesi e la mielopoiesi. È dalla linea mieloide che si sviluppano i globuli bianchi (granulociti e monociti), i megacariociti (responsabili della produzione di piastrine) e i globuli rossi. Dalla linea linfoide, invece, si differenziano per successivi stadi i diversi tipi di linfociti (B, T e natural killer). La cellula staminale pluripotente può seguire linee di maturazione morfologicamente e funzionalmente diverse, a seconda del condizionamento determinato dal tipo di stimolo o di mediatore presente. Questo dimostra l'incredibile plasticità e capacità di adattamento di queste cellule, fondamentali per la produzione di tutte le componenti del sangue necessarie per la vita.
La Transizione dell'Emoglobina: Un Indicatore della Maturazione
Un affascinante esempio della transizione e della maturazione del sistema emopoietico durante lo sviluppo fetale è rappresentato dalla progressiva sostituzione dei tipi di emoglobina. La transizione dall’eritropoiesi embrionale a quella fetale e infine a quella midollare è caratterizzata proprio dalla sostituzione dell’emoglobina embrionale con quella fetale (HbF) ed infine con l’emoglobina adulta (HbA2). Questo non è un semplice cambio, ma riflette un adattamento funzionale cruciale per la sopravvivenza e lo sviluppo del feto.
L'emoglobina embrionale, prodotta nelle prime fasi di sviluppo nel sacco vitellino, è ottimizzata per l'ambiente ipossico dell'utero precoce. Successivamente, l'emoglobina fetale (HbF), sintetizzata durante la fase epatica e poi midollare, possiede un'affinità per l'ossigeno significativamente più alta rispetto all'emoglobina adulta. Questa maggiore affinità è vitale per il feto, poiché gli permette di estrarre efficacemente l'ossigeno dal sangue materno attraverso la placenta, in un ambiente che è ancora relativamente ipossico rispetto a quello postnatale. Solo dopo la nascita, con l'avvio della respirazione polmonare e l'aumento della pressione parziale di ossigeno, l'emoglobina fetale viene gradualmente sostituita dall'emoglobina adulta (HbA), un processo che si completa nei primi mesi di vita extrauterina. Questa sequenza di sostituzioni non solo testimonia la capacità di adattamento dell'organismo, ma funge anche da marcatore biologico delle diverse fasi e delle sedi predominanti dell'emopoiesi, comprese le linee mieloidi che supportano la produzione degli eritrociti.
Regolazione e Dinamiche della Differenziazione Mieloide
L'impegno nella linea differenziativa, in particolare quella mieloide, non è un processo casuale, ma è finemente regolato. I progenitori ematopoietici e le cellule precursori vanno sempre di più sotto influenze regolatorie di fattori di crescita e ormoni. Ad esempio, per la produzione di globuli rossi, l'eritropoietina (EPO) è l'ormone regolatore. L'EPO è richiesta per il mantenimento di determinate cellule progenitrici eritroidi che, in assenza dell'ormone, vanno incontro alla morte cellulare programmata (apoptosi). Nel midollo osseo, il primo precursore eritroide riconoscibile morfologicamente è il pronormoblasto o proeritroblasto. Questa cellula può andare incontro a 4-5 divisioni cellulari che si risolvono nella produzione di 16-32 globuli rossi maturi. Con una produzione aumentata di EPO, il numero dei progenitori eritroidi è amplificato e a sua volta darà un aumento del numero degli eritrociti (globuli rossi). La regolazione della produzione di EPO è legata alla disponibilità di Ossigeno a livello del tessuto renale. L'organo responsabile della produzione di globuli rossi è chiamato eritrone.
Parallelamente alla linea eritroide, la mielopoiesi produce anche altre componenti essenziali del sangue. I globuli bianchi, come i granulociti neutrofili, vivono molto di meno e dopo sei ore sono destinati ad essere rimpiazzati da nuovi elementi. La produzione normale di globuli rossi deriva dalla giornaliera sostituzione del 0.8-1 % di tutti i globuli rossi circolanti nel corpo poiché la vita media di un globulo rosso è 100-120 giorni. Questi ritmi di produzione e ricambio sottolineano l'efficienza e la continua attività della mielopoiesi nel midollo osseo. Il sistema emopoietico è formato dagli organi che si incaricano dell’emopoiesi, cioè della generazione degli elementi cellulari del sangue: i globuli rossi, i globuli bianchi e le piastrine. Gli organi emopoietici formano il sistema tramite il quale l’organismo crea i globuli rossi, i globuli bianchi e le piastrine, quindi i cosiddetti elementi figurati del sangue. Il principale produttore di tali elementi è il midollo osseo, nello specifico quello localizzato all’interno dello sterno, delle vertebre, delle costole e delle scapole. Esso consente di rinnovare continuamente i globuli rossi, quelli bianchi e le piastrine (granulociti, linfociti e monociti) presenti nel sangue.
Inoltre, la ricerca di antigeni caratteristici di ciascuno stadio è un metodo molto usato negli studi sull’emopoiesi per comprendere meglio le dinamiche di differenziazione. Sebbene la dicotomia tra linea linfoide e mieloide sia stata a lungo accettata, basata su modelli murini, è stata recentemente messa in discussione a favore dell’ipotesi secondo cui la perdita di potenziale di maturazione avvenga in modo più graduale attraverso più cammini alternativi. Questa visione più sfumata riconosce la complessità della mielopoiesi, dove i percorsi di differenziazione potrebbero non essere sempre rigidamente binari, ma piuttosto un continuum di stati intermedi e di plasticità cellulare, che permette la generazione di monociti e globuli rossi dalla linea mieloide. Le scoperte che identificano popolazioni cellulari con potenziali di differenziazione specifici, come quelle che possono maturare pienamente lungo la linea linfoide ma produrre solo monociti (e non globuli rossi) lungo la linea mieloide, aprono nuove prospettive sulla comprensione dei primissimi stadi di indirizzamento delle staminali pluripotenti verso specifiche linee cellulari.
Siti di Emopoiesi Extra-Midollare: Meccanismi di Emergenza
Nonostante il midollo osseo sia la sede definitiva e primaria dell'emopoiesi, inclusa la mielopoiesi, l'organismo possiede meccanismi di riserva per far fronte a situazioni di stress o danno. Quando il midollo osseo è danneggiato o quando si verifica un’abnorme domanda di produzione di nuove cellule ematiche, la sua funzione può essere vicariata da siti di emopoiesi extra-midollare. I principali organi che possono riattivare questa funzione di emergenza sono il fegato e la milza. Questi organi, che in fasi precedenti dello sviluppo fetale erano essi stessi siti attivi di emopoiesi, possono ripristinare parzialmente questa capacità in condizioni di necessità.
Questa emopoiesi extra-midollare è un esempio della plasticità del sistema e della sua capacità di adattarsi a condizioni avverse, garantendo la produzione continua di cellule sanguigne, incluse quelle mieloidi, essenziali per la difesa e il trasporto di ossigeno. Nei casi più critici, dove la capacità rigenerativa endogena non è sufficiente, si ricorre a interventi medici come il trapianto di cellule staminali o di midollo osseo. In tali procedure, le cellule staminali emopoietiche vengono prelevate da un donatore e iniettate nel paziente che ne ha bisogno. Se donatore e paziente sono compatibili, le cellule infuse viaggiano fino al midollo osseo e iniziano la produzione di nuove cellule ematiche, ripristinando la mielopoiesi e l'emopoiesi complessiva.
Patologie e Malfunzionamento della Mielopoiesi
Il delicato equilibrio del processo emopoietico e mielopoietico può, purtroppo, essere compromesso, portando a diverse patologie. Esistono infatti numerose patologie legate al malfunzionamento dell’emopoiesi. Tra le più note e gravi si annoverano le Leucemie, in particolare le Leucemie mieloidi. Queste condizioni sono caratterizzate da una proliferazione abnorme e incontrollata di globuli bianchi immaturi derivanti dalla linea mieloide. In queste patologie, le cellule mieloidi immature, invece di maturare normalmente, si accumulano nel midollo osseo e nel sangue, interferendo con la produzione di altre cellule ematiche normali.
Il processo di differenziazione cellulare, che permette alle cellule di diventare sempre più "competenti" nel compiere una determinata funzione, può essere interrotto o alterato. Ad esempio, il globulo rosso maturo, con la sua forma di disco biconcavo e l'assenza di nucleo, è il risultato di una differenziazione che lo rende abile ad agganciare Ossigeno e Anidride carbonica all’emoglobina. Un malfunzionamento a questo livello può compromettere la funzionalità di intere linee cellulari, con conseguenze sistemiche per l'organismo. La comprensione delle sedi della mielopoiesi durante lo sviluppo fetale e delle sue intricate regolazioni è quindi fondamentale non solo per apprezzare la complessità della biologia umana, ma anche per diagnosticare e trattare efficacemente le patologie che derivano da un suo malfunzionamento.
tags: #sede #mielopoiesi #fetale