Introduzione all'Acetilcolina: Il Primo Neurotrasmettitore Scoperto
L'acetilcolina è un neurotrasmettitore fondamentale, una sostanza prodotta dal nostro organismo per trasferire gli impulsi nervosi in molteplici punti del sistema nervoso centrale e periferico. È stata il primo neurotrasmettitore ad essere scoperto, grazie agli studi di Otto Loewi, coronati nel 1924. Henry Hallett Dale e i suoi collaboratori, nel 1914, proposero per la prima volta la possibile azione di neurotrasmettitore da parte dell'acetilcolina. L'idea dell'esistenza di recettori come mediatori della trasmissione chimica degli impulsi era relativamente recente, essendo stata proposta la "sostanza ricettiva" da Langley nel 1905, proprio studiando l'azione di nicotina e curari sui recettori nicotinici della placca motrice. Successivamente, Otto Loewi definì il ruolo dell'acetilcolina nel sistema nervoso.
La sintesi dell'acetilcolina avviene nella parte pre-sinaptica dei neuroni, a partire da due molecole: l’acetil-CoA e la colina, entrambe coinvolte nei processi metabolici dell’organismo. In particolare, l’acetil-CoA interviene nel ciclo di Krebs aiutando a sintetizzare il citrato, mentre la colina interviene nella regolazione del colesterolo nel sangue. L'enzima che catalizza la reazione tra questi due reagenti per sintetizzare l'acetilcolina è la colinacetiltransferasi (ChAT). Trattandosi di una transferasi, il processo che media questo enzima prevede di unire il gruppo acetile dell’acetil-CoA alla colina. Questo avviene lungo il terminale assonale. Una volta sintetizzata, la molecola si immagazzina all'interno di vescicole. Al sopraggiungere di un impulso nervoso, le vescicole si legano alla membrana presinaptica, fondendosi e liberando per esocitosi il proprio contenuto nella fessura sinaptica, dove l'acetilcolina è libera di raggiungere i recettori postsinaptici e di interagire con essi.
Una volta che espleta il suo compito legandosi ai recettori del neurone post-sinaptico, intervenie un enzima chiamato acetilcolinesterasi (AChE). Dato che dal punto di vista molecolare l’ACh è un estere, per spezzare il legame è necessaria una molecola della classe delle esterasi. L’azione catalitica dell’AChE in presenza di acqua produce i reagenti di partenza, ossia acetil-CoA e colina. L’AChE si trova all’interno della giunzione sinaptica e, grazie all’idrolisi del neurotrasmettitore, permette che si possa trasmettere un impulso successivo. L'azione di questo enzima è molto importante, poiché consente di interrompere la trasmissione dell'impulso nervoso. La colina viene prontamente riassorbita dal terminale presinaptico e utilizzata per la sintesi di nuova acetilcolina. Pare che questo enzima si trovi anche sulla superficie di alcuni globuli rossi, anche se in questo caso non è del tutto nota la sua funzione.
I neuroni che secernono acetilcolina vengono definiti colinergici; analogo discorso per i suoi recettori. Sebbene l'acetilcolina sia un neurotrasmettitore eccitatorio, nell'ambito del sistema parasimpatico espleta azioni prevalentemente inibitorie. Questa molecola provoca, infatti, un rallentamento della frequenza cardiaca, mentre stimola la secrezione delle ghiandole bronchiali, salivari, gastriche e pancreatiche, aumentando la peristalsi intestinale e in genere tutte le funzioni digestive. L'acetilcolina si può riscontrare a livello delle sinapsi tra fibre pre-gangliari e neuroni post-gangliari del sistema nervoso simpatico e parasimpatico, e della midollare del surrene, così come in alcune sinapsi del sistema nervoso centrale.
I Recettori dell'Acetilcolina: Nicotinici e Muscarinici
I recettori dell'acetilcolina sono distinti in recettori nicotinici e muscarinici. Storicamente, la differenziazione delle due classi recettoriali è avvenuta utilizzando ligandi non endogeni: la muscarina per i recettori muscarinici e la nicotina per i recettori nicotinici. La nicotina è il ligando utilizzato per discriminare i recettori nicotinici da quelli muscarinici. Nonostante una diversità strutturale, l'acetilcolina è in grado di legarsi ad entrambi i recettori, poiché diversa è la parte di molecola che interagisce con i recettori muscarinici rispetto ai nicotinici. La diversa concentrazione e conformazione chimica di questi recettori, e delle relative isoforme nei tessuti, fa sì che i vari farmaci che interferiscono con l'azione dell'acetilcolina possano produrre effetti circoscritti prevalentemente ad un settore piuttosto che ad un altro.
I recettori di tipo muscarinico sono recettori metabotropici, associati quindi ad una proteina G. Nello specifico, i recettori M1, M3 e M5 sono accoppiati ad una proteina Gq/11 (proteina G di tipo eccitatorio) che promuove l’attivazione di un enzima, la fosfolipasi C (PLC), con conseguente aumento della concentrazione del calcio intracellulare. I recettori M1 si trovano soprattutto all’interno del sistema nervoso centrale (SNC) mentre gli M2 nel muscolo cardiaco.
Il Recettore Nicotinico dell'Acetilcolina (nAChR): Una Panoramica Generale
I recettori nicotinici (abbreviati in nAChR a causa del nome inglese Nicotinic acetylcholine receptors) sono recettori colinergici che formano canali ionici attivati da ligando nella membrana plasmatica di alcuni neuroni e nella porzione postsinaptica della giunzione neuromuscolare. In quanto recettori ionotropi, gli nAChR sono direttamente collegati al canale ionico e non fanno uso di secondi messaggeri, diversamente dai recettori metabotropici come i colinergici muscarinici.
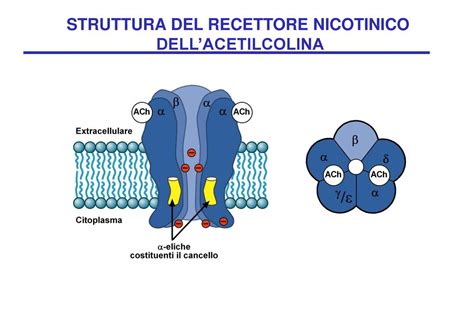
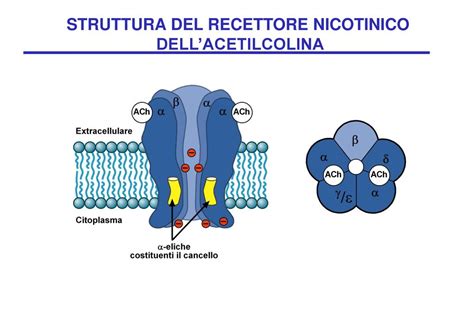
Il recettore colinergico nicotinico (nAChR) appartiene alla famiglia dei recettori-canale cationici nonché alla classe I (insieme al recettore serotoninico di tipo 3, al recettore del GABA di tipo A e al recettore glicinergico di tipo A). È un eteropentamero, ossia è costituito da cinque subunità proteiche che attraversano la membrana cellulare. Sono state clonate diverse subunità (alfa1-9, beta1-4, gamma, delta, epsilon) che possono assemblarsi tra loro a formare diverse isoforme che hanno proprietà biologiche e farmacologiche differenti. Per la loro struttura, quindi, i recettori nicotinici appartengono alla prima famiglia dei recettori ionotropi, detta dei recettori pentamerici o "Cys-loop".
Quando due molecole di acetilcolina si legano ai siti recettoriali presenti sulle subunità alfa, il recettore cambia leggermente la propria conformazione, aprendo un canale ionico. Questa apertura permette il passaggio all’interno della cellula di ioni positivi come sodio (Na+) e calcio (Ca++), e ioni potassio (K+) in uscita, con conseguente depolarizzazione ed eccitazione cellulare. Le funzioni dei recettori nicotinici sono principalmente la depolarizzazione della placca neuromuscolare in modo da permettere la contrazione.
Il recettore nicotinico è così chiamato perché ha alta affinità per il ligando non endogeno nicotina, media per l’acetilcolina e bassa per la muscarina.
Neuroscienze Di 2 Minuti: Nicotina
La Peculiarità del Recettore Nicotinico Muscolare Fetale
Nei vertebrati, i recettori nicotinici sono classificati grossolanamente in due sottotipi, in base al principale sito d'espressione: si distinguono, infatti, recettori nicotinici di tipo neuronale (NN) e di tipo muscolare (NM). Tra i recettori di tipo muscolare si hanno la forma embrionale e la forma adulta.
Considerando il recettore nicotinico muscolare nell’embrione, esso ha una struttura del tipo (2alfa1/1beta1/1gamma/1delta). La forma embrionale è composta dalle subunità α1, β1, δ, e γ in rapporto 2:1:1:1. Questa configurazione conferisce al recettore fetale delle proprietà ioniche e di apertura specifiche.
Nell’adulto, invece, la subunità gamma è sostituita da una subunità epsilon. Questa sostituzione modifica le proprietà ioniche del recettore. La subunità gamma è responsabile di una bassa conduttanza del recettore-canale, tanto che l’apertura del canale avviene in 11 ms. Nell’adulto, la conduttanza è alta e l’unica differenza riguarda l’espressione della subunità epsilon invece della gamma. In questo caso, il tempo di apertura raggiunge un massimo di 5 ms. Questa differenza nella velocità di apertura e nella conduttanza ionica riflette le diverse esigenze fisiologiche tra l'organismo in sviluppo e quello maturo.
Le subunità del recettore nicotinico muscolare sono disposte in maniera tale che una subunità alfa continui con la subunità gamma (o epsilon nell’adulto) e che l’altra subunità alfa confini con la subunità delta. Gli amminoacidi che partecipano al legame con l’ACh e altri farmaci sono localizzati nell’interfaccia tra la subunità alfa e la subunità gamma adiacente. Un’altra molecola di acetilcolina può riconoscere siti di legame dati da amminoacidi posti nell’interstizio tra la subunità alfa e la subunità delta. Nel sito di legame che riconosce la molecola di ACh è stato identificato un sito principale che lega la testa cationica dell’ACh (testa cationica ad azoto quaternario). Questo sito è formato da un amminoacido fondamentalmente presente sulla subunità alfa1; il legame con la testa cationica con tale amminoacido non fa altro che attirare amminoacidi carichi negativamente che sono posti sulla subunità gamma o sulla subunità delta.
Struttura Dettagliata del Recettore Nicotinico Muscolare
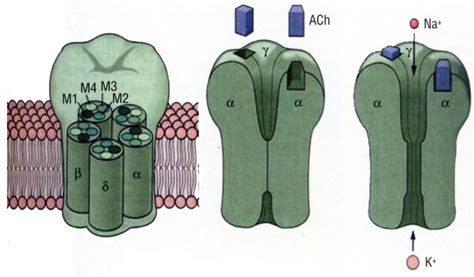
Come detto, i recettori nicotinici sono costituiti da 5 subunità, differenti a seconda del sottotipo recettoriale. Le subunità sono costituite da un'unica catena peptidica che, ripiegandosi su sé stessa, attraversa la membrana quattro volte. Vengono così identificati 4 domini transmembrana, nominati M1, M2, M3 e M4, uniti tra di loro a 3 anse (loop secondo la terminologia anglosassone), delle quali due intracellulari e una extracellulare. La seconda ansa intracellulare (quella che unisce M3 con M4) è costituita da una catena amminoacidica più lunga rispetto a quella delle altre due anse.
I domini amminoterminale (-NH2) e carbossiterminale (COOH) si trovano entrambi a livello extracellulare. In particolare, quello -NH2 è costituito da un numero maggiore di amminoacidi rispetto a quella -COOH terminale e racchiude la tasca necessaria per il legame con l'acetilcolina. La sezione extracellulare è la più voluminosa e sporge all'esterno della cellula di circa 60 Å. È in questa porzione che si trova il sito di legame per l'acetilcolina.
I domini M2 di ogni subunità sono quelli che circoscrivono il poro del recettore a livello della sezione transmembrana. Perciò, il poro, a livello della membrana, è delimitato solo da M2. La porzione transmembrana ha una lunghezza di circa 30 Å. Il poro del recettore, che ha forma a imbuto, tende a restringersi via via fino a raggiungere, nel punto più stretto, una larghezza di appena 7 Å. Poiché la membrana plasmatica è costituita da fosfolipidi (i quali hanno caratteristiche idrofobe), gli amminoacidi della sezione transmembranaria, per attraversarla, dovranno possedere spiccate caratteristiche lipofile. In questa sezione sono infatti riscontrabili un altissimo numero di amminoacidi lipofili che formano una catena peptidica che tende ad avvolgersi su sé stessa.
La sezione intracellulare è quella di minori dimensioni e ha una lunghezza di circa 20 Å. Sostanzialmente, ancora il recettore alla membrana e permette al Na+ di fluire all'interno della cellula. Poiché il citosol ha spiccate caratteristiche idrofile, anche gli amminoacidi presenti in tale sezione devono essere idrofili. La sezione intracellulare del recettore conta elevate quantità di amminoacidi idrofili che si dispongono nella specifica struttura secondaria definita foglietto β. Il passaggio da alfa elica a beta foglietto, che causa un restringimento nella struttura recettoriale, è dovuto a una molecola di prolina che non è in grado di formare ponti idrogeno intramolecolari, consentendo il passaggio alla struttura a beta foglietto.
Sezionando trasversalmente il canale, si nota che questo da una serie di anelli concentrici nei quali gli amminoacidi vengono ripetuti. Un anello interno, che contiene aspartato (D 234), carico negativamente, attira i cationi nel canale. I tre anelli idrofobici sono necessari per eliminare l'acqua che solvata gli ioni. Gli ioni in ambiente fisiologico sono idratati, e perciò sono molto voluminosi e non riuscirebbero ad attraversare il canale, che risulterebbe troppo stretto. Per poter entrare, devono perdere l'acqua che li circonda: gli amminoacidi di questi tre anelli, essendo idrofobici, respingono l'acqua all'esterno consentendo solo ai cationi di entrare.
Meccanismo di Legame dell'Acetilcolina e Apertura del Canale
Indipendentemente dalle strutture recettoriali, l'unica subunità in grado di legare l'acetilcolina è la subunità α, che presenta la specifica tasca all'interno della quale il ligando si può collocare. Questo significa che, per essere attivato, il recettore della placca neuromuscolare (che presenta 2 subunità α) deve legare contemporaneamente 2 molecole di acetilcolina, una per ogni subunità α. Per il recettore del SNC, che invece presenta 5 subunità di tipo α7, sono necessarie 5 molecole di acetilcolina per poter trasdurre il segnale.
Il sito di legame per l'acetilcolina si trova all'interfaccia tra la subunità alfa e la subunità non-alfa (causa la variabilità delle forme recettoriali, le subunità beta, gamma, delta ed epsilon vengono definite per comodità non-alfa). Il sito che lega l'acetilcolina è perciò costituito da una cavità fra la subunità alfa e non alfa. La cavità funzionalmente più importante è a livello della subunità alfa, definito sito principale, e contribuisce in maniera rilevante al legame con il ligando e alla modificazione conformazionale del recettore, mentre la cavità formata dalle subunità gamma o delta viene definita sito complementare. Le tre anse della subunità alfa vengono indicate con le lettere A, B e C, mentre quelle sulla subunità non-alfa sono nominate D, E e F.
Nel sito di legame che riconosce la molecola di ACh, è stato identificato un sito principale che lega la testa cationica dell’ACh, ad azoto quaternario. Questo sito è formato da un amminoacido fondamentalmente presente sulla subunità alfa1. Il legame con la testa cationica non fa altro che attirare amminoacidi carichi negativamente che sono posti sulla subunità gamma o sulla subunità delta. Amminoacidi importanti per il legame includono quelli che formano un ponte disolfuro giallo, due tirosine, una treonina e un triptofano.
In ultima analisi, si crea un avvicinamento di queste subunità tra di loro, determinando uno slittamento di α-eliche transmembrana responsabili dell’aumento della dimensione del poro-canale. A seguito dell'interazione con l'acetilcolina, il recettore cambia la propria conformazione, permettendo l'apertura del canale. Poiché la selettività nei confronti dei cationi non è assoluta, ioni di medesima carica e di raggio atomico simile possono fluire attraverso il canale. Viene permesso perciò il flusso in particolare di ioni Na+, ma anche di Ca++ (entrambe in entrata), e di K+ in uscita.
Recettori Nicotinici Neuronali (NN): Localizzazione e Funzioni
I recettori nicotinici neuronali (NN) hanno sede nel SNC, nei gangli del sistema nervoso autonomo (SNA) e nella midollare del surrene. Le loro funzioni, quando legano l’ACh o agonisti, sono molteplici. Nel SNC, sono coinvolti nei meccanismi dell’attenzione, con un ruolo che è ancora oggetto di studio e non è del tutto definito. A livello dei gangli, determinano la depolarizzazione dei neuroni postgangliari, partecipando così alla trasmissione gangliare. Infine, nella midollare del surrene, contribuiscono a determinare la secrezione di catecolamine, come adrenalina, dopamina e noradrenalina.
Il recettore nicotinico del SNC ha una struttura del tipo 2alfa/3beta. Sono espresse tutte le isoforme tranne quelle espresse nel recettore nicotinico muscolare; infatti, le isoforme alfa possono essere presenti dalla 2 alla 10, tranne la 1 che è tipica del recettore muscolare, come le subunità beta che sono espresse in tutte le isoforme, da 2 a 4, tranne la 1 tipica del muscolare. Il recettore ad alta densità, formato dall’isoforma alfa4 e dalla beta2 (2alfa4/3beta2), come il recettore nicotinico di placca, riconosce ed è sensibile alla nicotina ma non lega l’α-bungarotossina, che è una tossina paralizzante.
A livello del SNC ci sono altri due tipi di recettore formati da 5 subunità uguali (5alfa7 o 5alfa9) la cui presenza conferisce a questo recettore un’elevata permeabilità al calcio. Questa è una proprietà controcorrente, dato che i recettori nicotinici muscolari hanno una spiccata permeabilità al sodio. Tali recettori, però, sono sensibili all’α-bungarotossina.
Farmacologia dei Recettori Colinergici: Agonisti e Antagonisti
Visto il ruolo fondamentale dell’acetilcolina, è facile intuire quali danni potrebbe portare assumere sostanze che ne bloccano o alterano l’azione di trasmissione. Gli antagonisti nicotinici sono farmaci anticolinergici che agiscono a livello dei recettori per l'acetilcolina di tipo nicotinico (neuronale e muscolare). Essi trovano impiego, soprattutto, come rilassanti dei muscoli. I farmaci anticolinergici impediscono all'acetilcolina di legarsi al suo recettore (nicotinico o muscarinico), presente nelle cellule nervose, inibendo gli impulsi del sistema nervoso parasimpatico. Tali impulsi nervosi sono responsabili dei movimenti muscolari involontari nel tratto gastrointestinale, nel polmone, nel tratto urinario e in altre parti del corpo. In funzione della loro tipologia (muscarinici o nicotinici), il campo di impiego dei farmaci anticolinergici è molto vasto.
Possiamo distinguere tra farmaci e veleni che agiscono come agonisti o antagonisti. Le molecole che interagiscono con i recettori possono avere un comportamento agonista od antagonista.
Agonisti e Antagonisti Muscarinici
Gli antagonisti muscarinici sono farmaci anticolinergici che esplicano la loro attività interagendo con i recettori dell'acetilcolina di tipo muscarinico. Ne è un esempio la scopolamina, un farmaco utilizzato per alleviare nausea e capogiri ed evitare il vomito in caso di mal d'auto o alla ripresa di conoscenza dopo un'anestesia. Il farmaco atropina blocca i recettori muscarinici, inibendo l'attività del parasimpatico (parasimpaticolitico).
La pilocarpina, un farmaco utilizzato soprattutto in oftalmologia per restringere la pupilla e stimolare la lacrimazione dell'occhio (utile nel trattamento del glaucoma), è un agonista muscarinico; si lega, infatti, ai recettori muscarinici dell'acetilcolina. In tal senso, la pilocarpina contrasta l'azione dell'atropina.
Agonisti e Antagonisti Nicotinici
Il curaro, un estratto vegetale velenoso, blocca l’azione dell’ACh prima che si leghi ai recettori nicotinici muscolari. A livello fisiologico, il curaro provoca morte per paralisi muscolare, bloccando l'azione dell'acetilcolina sulle membrane muscolari (dove si trovano recettori nicotinici). Questa condizione parte a livello del collo per poi scendere lungo gli arti e verso l’addome fino a che non si paralizza il diaframma, impedendo la respirazione.
Esistono anche i bloccanti depolarizzanti, come la succinilcolina che, attraverso un legame irreversibile, desensibilizzano il recettore nicotinico, provocando paralisi muscolare e respiratoria.
Sostanze che Modulano l'Acetilcolinesterasi (AChE)
Una sostanza che, al contrario del curaro, ha un effetto contrario sull'acetilcolina è la fisostigmina, contenuta nella fava del Calabar. Si tratta sempre di un alcaloide, ma prolunga l’azione del neurotrasmettitore bloccando la colinesterasi. Si utilizza come farmaco, ma se dato in dosi eccessive può provocare convulsioni, bradicardia e arrivare a causare un arresto cardiaco.
Anche i gas nervini bloccano questo enzima, facendo sì che l'acetilcolina rimanga ancorata ai suoi recettori. L'effetto letale di questi gas ci è utile per indagare gli effetti dell'interazione tra acetilcolina e i suoi recettori muscarinici: tosse, oppressione toracica, ipersecrezione bronchiale fino all'edema polmonare, nausea, vomito, diarrea, aumento della salivazione, miosi e difficoltà nella visione, riduzione della frequenza cardiaca fino all'arresto e incontinenza urinaria. A causa dell'accumulo di acetilcolina nei recettori nicotinici, insorgono invece sintomi come: pallore cutaneo, tachicardia, ipertensione arteriosa, iperglicemia e alterazioni a carico del sistema muscolo-scheletrico, in particolare astenia e facile esauribilità muscolare, tremori e crampi. Per l'accumulo di acetilcolina, la muscolatura scheletrica può essere paralizzata e si può avere morte per paralisi muscolare in contrazione. Infine, gli effetti sul sistema nervoso centrale includono contrazioni tonico-cloniche di tipo epilettiforme, fino alla depressione dei centri respiratori ed alla morte per paralisi del diaframma e dei muscoli intercostali.
Sostanze che Influenzano il Rilascio di Acetilcolina
Anche il veleno della vedova nera agisce sulla trasmissione chimica dell’impulso nervoso, portando a un eccesso di rilascio di acetilcolina. Questo porta a una condizione di tetania nei muscoli, che rimangono in contrazione senza potersi rilassare.
Il botulino, una tossina velenosissima utilizzata in concentrazioni infinitesimali in medicina estetica, ha a che fare con l'acetilcolina; con la sua azione, infatti, ne impedisce il rilascio dalle vescicole. In questo modo, il botulino causa una paralisi flaccida dei muscoli, divenendo mortale quando coinvolge pesantemente quelli respiratori; si contrappone, in tal senso, all'azione del tetano, caratterizzata da paralisi spastica che risulta tuttavia indipendente dall'acetilcolina.
Siti di Modulazione del Recettore Nicotinico
Il recettore colinergico, oltre al sito principale che riconosce l’ACh, gli agonisti selettivi come la nicotina e gli antagonisti competitivi specifici che competono con l’ACh per legarsi sullo stesso recettore, presenta anche dei siti di modulazione. Tra questi ce ne sono due che legano farmaci e sono i siti per i glucocorticoidi. I glucocorticoidi sono usati anche nelle malattie autoimmunitarie, tra cui la miastenia grave. È importante conoscere l’esistenza di questo sito di modulazione del recettore nicotinico per gli steroidi perché, attraverso tale legame, gli steroidi inibiscono l’attività del recettore nicotinico e sono importanti per comprendere gli effetti secondari dei glucocorticoidi a livello dei recettori nicotinici nei muscoli respiratori.
Nel trattamento della miastenia grave si utilizzano dei farmaci inibitori dell’ACh-esterasi che potenziano la trasmissione colinergica, la quale è carente in questa malattia. Tra questi c’è l’eserina, il cui effetto benefico è dovuto a un duplice meccanismo: inibire l’ACh-esterasi, mantenendo un buon tenore di ACh e potenziando la trasmissione colinergica; e legare direttamente il recettore nicotinico in un sito di modulazione allosterica, favorendo l’attività del recettore stesso.
I Neonicotinoidi: Agonisti Selettivi del Recettore Nicotinico
I neonicotinoidi, inventati dal laboratorio Shell di Modesto in California nei primi anni '70 ed immessi sul mercato nel 1991, oggi coprono un fatturato dell’ordine di un paio di miliardi di dollari, circa un sesto del mercato mondiale degli insetticidi. Il termine neonicotinoide fu proposta per la prima volta da I. Yamamoto. Un collega faceva notare tempo fa che non capiva come mai i neonicotinoidi si chiamassero così visto che non assomigliano poi molto alla nicotina.
I neonicotinoidi, definiti così poiché sono dei nuovi agonisti del recettore dell’acetilcolina di tipo nicotinico, si legano ai recettori stessi e ne causano una sovrastimolazione. Inoltre, essi non vengono distrutti dall’azione dell’enzima acetilcolinesterasi, deputato a distruggere l’acetilcolina e a impedire i danni da sovrastimolazione. In questo modo, i neonicotinoidi sono in grado di sovrastimolare i recettori in modo irreversibile, portando a morte e a paralisi gli organismi. Le basi di tale selettività sono squisitamente chimiche e si aggiungono alle differenze nella composizione dettagliata del recettore fra le varie specie e nei vari tessuti della medesima specie.
L’acetilcolina è rappresentata comunemente come ione positivo, poiché la forma attiva della molecola è proprio questo ione. Tutti i suoi agonisti che manchino di una carica positiva in particolare sull’azoto pirrolico (o in posizione analoga) non possono quindi avere effetti altrettanto efficaci. La stessa nicotina, in condizioni di pH fisiologico (7.4), ha l’azoto pirrolico carico positivamente; questa carica positiva dà a questi ioni una forte affinità per i recettori acetilcolinici-nicotinici dei mammiferi. D’altronde, questa stessa carica rende tale molecola un insetticida efficace ma non particolarmente forte, in quanto per agire deve raggiungere i recettori acetilcolinici che negli insetti si trovano solo a livello di sistema nervoso centrale, mentre invece nei mammiferi essi si trovano anche nel sistema nervoso periferico, poiché controllano funzioni cruciali come la respirazione. La conclusione è quindi che, mentre la nicotina che ha una carica netta esercita i suoi effetti intossicanti più efficacemente sul sistema nervoso periferico dei mammiferi (per esempio nei polmoni dei fumatori), essa non è in grado di attraversare che parzialmente la barriera ematoencefalica degli insetti ed agire sul loro sistema nervoso centrale e quindi non è un insetticida altrettanto potente.
I neonicotinoidi non possiedono un azoto carico in posizione opportuna a livello del pH umano e quindi non sono carichi ed agiscono meno efficacemente sui recettori acetilcolinici umani; questo spiega la selettività dei neonicotinoidi, che si polarizzano ma non hanno carica netta nella posizione opportuna. In conclusione, ma senza alcuna idea di rigore assoluto, i neonicotinoidi agiscono bene non perché assomigliano molto alla nicotina, come potrebbe far pensare il loro nome, ma proprio perché non assomigliano completamente alla nicotina: non si caricano, ma proprio per questo costituiscono un buon strumento per superare la barriera ematoencefalica degli insetti che rallenta invece la nicotina come insetticida.
I neonicotinoidi costituiscono un buon esempio di come la chimica possa comprendere ed agire sui meccanismi più intimi della vita; proprio per questo il loro uso indiscriminato può avere potenti effetti su quegli insetti che, attraverso il meccanismo dell’impollinazione, garantiscono la fertilità di buona parte della nostra agricoltura (non solo le api, quindi, ma anche i bombi per esempio e tutti gli altri impollinatori), un servizio economico per cui non sono pagati. La sospensione, sia pur parziale e temporanea, dei neonicotinoidi è al momento l’unica iniziativa ragionevole che possa salvare questi preziosissimi collaboratori agricoli.
Metodi di Studio dei Recettori Nicotinici
Lo studio dei recettori nicotinici è stato lungo e continua attualmente, con lo scopo di migliorarne le informazioni e identificare ligandi selettivi da poter utilizzare in terapia. La ricerca, con il passare degli anni, si è affinata, arricchendosi di nuove tecniche.
Mutagenesi
La mutagenesi è una tecnica che prevede la modificazione di un amminoacido strutturale del recettore. Variando tale amminoacido, si ottiene un recettore con caratteristiche diverse da quello selvaggio. Se ne studiano le differenze e se ne valutano le nuove caratteristiche. Per esempio, sostituendo nella regione di legame del ligando l'amminoacido aspartato con la valina, il binding dell'acetilcolina avviene con maggiore difficoltà.
Una variante di questa metodica è la mutagenesi a doppio scambio: si muta un amminoacido con un altro, e nel nuovo amminoacido introdotto viene variato il gruppo funzionale. Per esempio, l'aspartato (che possiede un -COOH) viene scambiato con una lisina (amminoacido basico), introducendo in quest'ultima un gruppo carbossilico.
Cristallografia
La cristallografia è una tecnica di indagine che permette lo studio del recettore ai raggi X. È una tecnica importante nello studio degli enzimi, ma che risulta molto più complicata quando applicata ai recettori. Infatti, i recettori molto difficilmente tendono a cristallizzare, poiché precipitano in forma amorfa la quale non consente lo studio con la tecnica cristallografica. Attualmente il recettore nicotinico non è stato ancora cristallizzato, perciò non è stato possibile studiarlo mediante i raggi X.
Proteine Affini
Ai fini dello studio della funzionalità strutturale, è stata importantissima una scoperta, da parte di un gruppo di ricercatori olandesi, di una particolare proteina affine strutturalmente al recettore nicotinico. Tali ricercatori hanno infatti isolato, da alcune lumachine di mare, una proteina, definita Ach binding protein (proteina legante l'acetilcolina), che presenta una affinità di struttura pari al 40% di quella del recettore nicotinico. Questa è una proteina citosolica che ha la funzione di legare l'acetilcolina, in modo da intrappolarla e porre fine all'attività dell'acetilcolina stessa. Svolge perciò, nelle lumachine di mare, un ruolo similare a quello dell'acetilcolinesterasi nell'uomo.
tags: #recettore #fetale #acetilcolina