La riproduzione è un processo fondamentale per la sopravvivenza di ogni organismo vivente, comprese le piante. Uno dei modi principali attraverso cui le piante generano prole è la produzione di semi, e l'impollinazione è il passaggio cruciale che assicura che possa aver luogo il processo di fecondazione e la conseguente formazione dei semi. Tra le diverse strategie adottate dalle piante per garantire la propria propagazione, l'autoimpollinazione rappresenta un meccanismo affascinante e particolarmente efficace in determinate condizioni.
Il Meccanismo dell'Autoimpollinazione: Autogamia e Geitonogamia
L'autoimpollinazione, conosciuta anche come autogamia, si verifica quando il polline passa direttamente dall'antera di un fiore allo stigma dello stesso fiore (questo è il caso dell'autogamia in senso stretto) o di fiori dello stesso individuo (un fenomeno noto come geitonogamia). Le piante che praticano questo tipo di impollinazione sono dette autogame. Questo processo è possibile solo nei fiori ermafroditi, ossia quelli che possiedono sia organi riproduttivi maschili (stami con antere) che femminili (pistillo con stigma).
In alcune specie, l'autoimpollinazione è addirittura obbligata, come nelle piante cleistogame, che producono fiori che non si schiudono mai, garantendo così che il polline delle antere raggiunga lo stigma senza alcuna interferenza esterna. Un esempio notevole della capacità delle piante di adattarsi alle condizioni ambientali e ai cambiamenti negli impollinatori è emerso da uno studio condotto su piante di rapa (Brassica rapa). Questo esperimento controllato ha mostrato come le piante esposte a impollinatori meno efficienti, come i sirfidi (Episyrphus balteatus), abbiano sviluppato una più efficiente autoimpollinazione. Questo dimostra che un'alterazione nella composizione degli insetti impollinatori non si ripercuote solo sul funzionamento dell'ecosistema, ma anche sull'evoluzione delle caratteristiche dei vegetali, ivi compreso il sistema di accoppiamento usato.
Vantaggi e Svantaggi dell'Autoimpollinazione: Equilibri Genetici e Adattamenti
Sebbene l'autoimpollinazione garantisca una riproduzione certa anche in assenza di impollinatori esterni o in ambienti dove questi sono scarsi, è generalmente considerata meno vantaggiosa dell'impollinazione incrociata. L'impollinazione incrociata, infatti, favorisce la comparsa di nuove combinazioni genetiche, aumentando la diversità genetica della popolazione e, di conseguenza, la sua capacità di adattarsi a cambiamenti ambientali e di resistere a malattie.
Nonostante questo potenziale svantaggio in termini di diversità genetica, l'autoimpollinazione è frequente nelle piante annue a vita breve, dove la rapidità e l'efficacia della riproduzione sono prioritari per assicurare la sopravvivenza della specie in un ciclo vitale accelerato. Questo meccanismo offre una soluzione affidabile per garantire la produzione di semi, anche quando le opportunità di impollinazione incrociata sono limitate.
Barriere all'Autoimpollinazione: Strategie per Favorire la Diversità Genetica
Nella maggior parte dei casi, esistono adattamenti nelle piante per evitare l'autoimpollinazione, proprio per promuovere la diversità genetica offerta dall'impollinazione incrociata. Questi adattamenti possono essere di natura fisica o chimica.
Uno dei meccanismi per prevenire l'autoimpollinazione è la produzione di fiori unisessuati su individui di sesso diverso, come avviene nelle specie dioiche. In queste piante, gli individui sono distinti in maschili e femminili, rendendo impossibile l'autoimpollinazione e obbligando all'impollinazione incrociata.
Nelle specie con fiori ermafroditi e in quelle monoiche, dove fiori maschili e femminili si trovano sulla stessa pianta, sono noti altri meccanismi per favorire l'incrocio. Un meccanismo efficace è l'autoincompatibilità (o autosterilità), cioè l'impossibilità per una pianta bisessuale di produrre zigoti con il suo stesso polline. In pratica, il polline non può germinare sullo stigma dello stesso individuo, e se germina, non feconda gli ovuli. L'autoincompatibilità è geneticamente controllata in molte specie da alleli multipli di un gene chiamato S, che impediscono al polline geneticamente simile di completare il processo di fecondazione. Ad esempio, nel melo, si ha l'autoincompatibilità: per avere una regolare fecondazione e il conseguente sviluppo del frutto occorre prevedere la fecondazione incrociata. Questo significa che due varietà diverse ma compatibili devono essere piantate vicine per garantire una buona produzione di frutti.

L'Impollinazione Incrociata (Allogamia): Il Ruolo della Diversità
L'impollinazione incrociata, o allogamia, si verifica quando il polline viene trasportato dall'antera di un fiore allo stigma del fiore di un individuo differente della stessa specie. Meno frequentemente, può avvenire anche tra fiori di specie diverse, originando ibridi. Le piante che praticano questo tipo di impollinazione sono dette allogame e rappresentano la maggioranza delle piante a fiore. Questo processo è fondamentale per mantenere la vitalità genetica delle popolazioni vegetali.
I Fondamentali dell'Impollinazione: Trasporto del Polline e Gamia
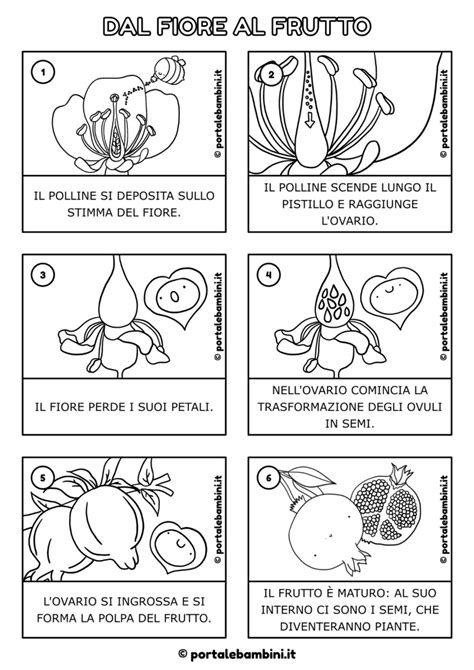
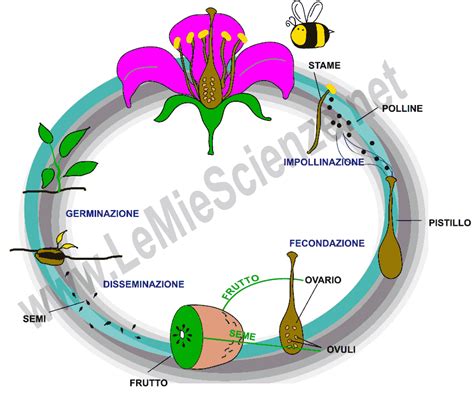
Per comprendere appieno l'autoimpollinazione e l'impollinazione incrociata, è essenziale partire dalla definizione generale del processo. L'impollinazione è il trasporto dei granelli di polline dalla parte maschile a quella femminile dell'apparato riproduttivo delle piante che producono semi (Spermatofite). Il polline è contenuto nelle sacche polliniche dei coni delle Gimnosperme o negli stami dei fiori delle Angiosperme. L'impollinazione consente la successiva gamia (unione di gameti maschili e femminili) e la produzione dei semi, che rappresentano la prole per la prossima generazione.
La germinazione del polline è un processo complesso che ha tre fasi distinte: idratazione, attivazione ed estensione del tubetto pollinico. La germinazione avviene solo dopo la reidratazione, un meccanismo che impedisce che la germinazione avvenga prematuramente nell'antera. L'idratazione consente alla membrana plasmatica del granulo pollinico di riformarsi nella sua normale organizzazione a doppio strato, fornendo un'efficace membrana osmotica essenziale per il processo. Successivamente, l'attivazione del polline comporta a livello cellulare lo sviluppo di filamenti di actina in tutto il citoplasma della cellula, che alla fine si concentrano nel punto da cui emergerà il tubo pollinico, il quale guiderà i gameti maschili verso l'ovulo. La germinazione del polline è molto difficile in ambiente artificiale, in quanto la natura ha evoluto meccanismi per garantire che avvenga solo sullo stigma della pianta corretta.
Gli Agenti Impollinatori: Vettori Biotici e Abiotici
Il trasporto del polline, sia nell'autoimpollinazione che nell'impollinazione incrociata, può essere attuato da diversi vettori, sia biotici (organismi viventi) che abiotici (fattori non viventi). L'impollinazione biotica si basa su impollinatori viventi che spostano il polline da un fiore all'altro, mentre l'impollinazione abiotica si basa su vento o acqua.
L'impollinazione abiotica, utilizzando metodi non viventi, consente alla pianta di spendere energia direttamente sul polline piuttosto che nell'attirare gli impollinatori con fiori colorati o odorosi e nettare, risorse che richiedono un notevole investimento energetico.
Impollinazione Abiotica: Vento e Acqua
L'impollinazione anemogama, o anemofila, è un tipo di impollinazione abiotica che utilizza come mezzo di dispersione il vento. Non essendoci l'intervento di animali, le piante anemofile non producono nettare o comunque non presentano adattamenti per attirare i pronubi (impollinatori). Tuttavia, l'impollinazione anemofila può comunque determinare degli adattamenti morfologici per disperdere e/o attrarre al meglio i granuli pollinici. Gli stami possono presentare filamenti allungati, esponendo a maturità le antere al di fuori del fiore per facilitare la dispersione del polline. Allo stesso modo, il gineceo può presentare uno stigma piumoso per meglio intercettare il polline aerodisperso. Esempi di granuli pollinici aerodispersi in sud Europa includono quelli di Plantago sp., Urticaceae (Paretaria sp.), Castanea sp., Quercus sp., Myrtaceae (Callistemon sp.), Platanus sp., Poaceae e Oleaceae, nonché Cupressus sp.
L'impollinazione idrogama, o idrofila, è un metodo di trasporto del polline meno diffuso in cui il polline è distribuito dal flusso delle acque, in particolare nei fiumi e nei torrenti. Un esempio interessante è rappresentato da alcune piante acquatiche come Ceratophyllum o Vallisneria. In quest'ultima specie, i fiori maschili, giunti a maturazione, si staccano dalla pianta che li ha prodotti e vengono trasportati dalle correnti fino a contatto con i fiori femminili. Altre modalità di impollinazione idrogama comprendono la distribuzione del polline in ambiente completamente sommerso.
Impollinazione Biotica (Zoogama): La Collaborazione Animale-Vegetale
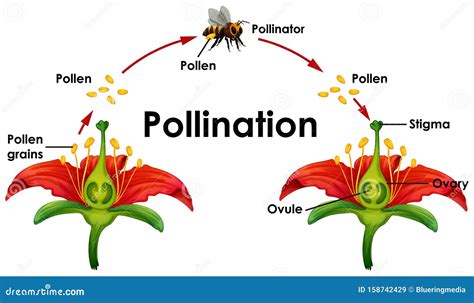
Più comunemente, l'impollinazione coinvolge gli impollinatori (detti anche pronubi): organismi che trasportano o spostano i granuli pollinici dall'antera di un fiore alla parte ricettiva del carpello o del pistillo (stigma) di un altro. Tra le 100.000 e le 200.000 specie di animali agiscono come impollinatori delle 250.000 specie di angiosperme del mondo. La maggior parte di questi impollinatori sono insetti, ma circa 1.500 specie di uccelli e mammiferi visitano i fiori e possono trasferire polline tra di loro. Anche rettili, piccoli marsupiali, mammiferi e molluschi possono svolgere questa funzione.
La coevoluzione di piante e animali impollinatori ha portato in molti casi ad una strettissima interdipendenza. Le specie entomofile (impollinate dagli insetti) possono presentare specifici adattamenti volti ad attirare con maggiore efficacia gli insetti impollinatori, come ad esempio il colore, il profumo e la produzione di nettare. Le piante inducono gli animali ad impollinarle offrendo loro un premio oppure, in alcuni casi, ingannandoli. Gli impollinatori non visitano i fiori con lo scopo altruistico di fornire il servizio di impollinazione; normalmente raccolgono nettare e/o polline e nel far questo si ricoprono di polline che viene così trasportato da una pianta all'altra, permettendone la riproduzione. Le specie che svolgono funzione impollinatrice sono diverse, e tra queste spiccano gli insetti, i bombi, le api (sia selvatiche che allevate), ma anche animali vertebrati.

Forme Specifiche di Impollinazione Animale
L'impollinazione entomogama, o entomofila, è il processo di impollinazione portato avanti da insetti, che rappresentano la categoria più numerosa e diversificata di impollinatori. In agricoltura, gli impollinatori, come i bombi o le api allevate, promuovono una più ricca e migliore produzione e in certi casi la rendono possibile; senza di essi non avremmo ciliegie, mele, meloni e zucchine.
L'impollinazione ornitogama è quella in cui il vettore dell'impollinazione è rappresentato dagli uccelli. I fiori impollinati dagli uccelli hanno solitamente una colorazione molto vistosa (spesso gialla o rossa) e una abbondante produzione di nettare. Gli uccelli coinvolti in tale processo sono specie nettarivore, ossia si nutrono del nettare prodotto dai fiori introducendo il lungo becco nel calice del fiore. Tra di essi vi sono i colibrì (Trochilidae), gli honeyeaters australiani (Meliphagidae), le nettarinie africane (Nectariniidae) ed alcune specie di pappagalli della famiglia Psittacidae.

Nell'impollinazione chirotterogama, il vettore dell'impollinazione è costituito dai pipistrelli. Tale modalità è comune tra le piante i cui fiori si schiudono durante la notte, spesso con caratteristiche che attraggono questi impollinatori notturni. Tra di esse si annovera il baobab Adansonia digitata ed altre specie della famiglia Bombacaceae.
Specie che svolgono funzione impollinatrice sono state descritte anche tra i sauri, in particolare tra le lucertole (per esempio Podarcis lilfordi), i gechi (per esempio il geco mauriziano Phelsuma ornata) e gli scinchi (per esempio Leiolopisma telfairii). Questo fenomeno è particolarmente interessante e spesso si manifesta come un adattamento insulare.
È interessante notare che l'impollinazione era stata già descritta da Filippo Buonanni della Compagnia di Gesù (1638-1725), prefetto della biblioteca del Collegio Romano e direttore del Museo Kircheriano, che nel 1691 aveva pubblicato un libro in cui polemizzava con Redi, sostenendo la generazione spontanea, ma comunque accennando al processo.
L'Importanza Cruciale dell'Impollinazione per Ecosistemi e Umanità
L'impollinazione da parte degli animali rappresenta un servizio ecosistemico di grande valore per l'umanità, sia dal punto di vista economico che ecologico. Senza gli impollinatori, l'uomo non sopravvivrebbe e gli ecosistemi terrestri collasserebbero. Se l'impollinazione zoogama non avesse luogo, l'85% delle piante a fiore non potrebbe riprodursi per via sessuale. Molte tra esse sono specie coltivate indispensabili per l'alimentazione umana. Delle 115 piante maggiormente coltivate in tutto il mondo, il 75% richiede l'impollinazione da parte degli animali, costituendo il 35% della resa delle colture a livello mondiale. Per questo motivo in taluni casi, nelle coltivazioni intensive quando l'impollinazione è scarsa si deve ricorrere all'azione diretta dell'uomo tramite l'impollinazione artificiale.
Esiste un ulteriore aspetto che rende l'impollinazione mediata dagli animali un processo cruciale, e riguarda la spinta evolutiva che questo ha dato alla diversificazione di piante a fiore e animali, in particolar modo insetti. L'85% delle piante a fiore affida la propria impollinazione agli animali e il 65%, in particolare, agli insetti. La produzione di sostanze di "ricompensa", quali polline e nettare, e lo sviluppo di strutture fiorali attrattive, ha reso possibile l'instaurarsi di relazioni mutualistiche tra specie vegetali e impollinatori.
43 Radio Veneto Agricoltura - Impollinatori e biodiversità: il forte legame con l’agricoltura
L'Intervento Umano: Impollinazione Artificiale e Gestione Agronomica
L'impollinazione artificiale, detta anche manuale o meccanica, è una tecnica utilizzata in agricoltura quando l'impollinazione spontanea è insufficiente. Avviene grazie al trasferimento manuale del polline per mezzo di appositi pennelli o direttamente strofinando i fiori maschili sugli stigmi dei fiori femminili. Il ricorso a tali tecniche può essere reso necessario dalla mancanza di impollinatori naturali, dovuta alla coltivazione di specie al di fuori della loro area di origine (come per esempio nel caso della Vanilla planifolia, che nel suo ambiente naturale è impollinata da una specifica specie di ape), oppure può essere legata a necessità di selezione di una specifica varietà, o ancora può essere un modo per supplire alla carenza di impollinatori conseguente all'uso intensivo di fitofarmaci, una delle cause della Sindrome dello spopolamento degli alveari.
L'impatto degli impollinatori e l'efficacia dell'impollinazione sono temi complessi. Ad esempio, è stata sollevata la domanda su quanto incida sul frutto la pianta impollinatrice quando si devono impollinare tra loro due varietà di piante autosterili (es. nespolo, albicocco). Sembra che, in generale, la pianta impollinatrice non trasmetta i suoi caratteri ai frutti della pianta impollinata. Questa osservazione si basa sul fatto che, ad esempio, un ciliegio "durona" continua a produrre le sue caratteristiche ciliegie anche se attorniato da ciliegi selvatici e ciliegi acidi, e le diverse varietà di zucchine producono sempre i propri frutti, pur impollinandosi vicendevolmente. Si crede che ciò dipenda dal fatto che il fiore sboccia sul frutticino già formato che ha le sue caratteristiche genetiche predeterminate. Diverso è il discorso se si usa il seme per ottenere nuove piante, dove la combinazione genetica è evidente. Tuttavia, è stato notato che i frutti di nespolo e albicocco possono variare parecchio da un anno all'altro, suggerendo una possibile influenza indiretta dell'impollinazione.
Negli studi sull'evoluzione delle piante in risposta a cambiamenti negli impollinatori, è stato dimostrato che le alterazioni nella composizione degli impollinatori possono influenzare l'evoluzione delle caratteristiche vegetali. Nell'esperimento sulla Brassica rapa, le piante impollinate dai bombi si sono evolute in modo da essere più alte, avere fiori più profumati (con l'emissione di una quantità quasi doppia di sostanze volatili) e petali con una maggiore capacità di riflettere la luce nello spettro dell'ultravioletto, pur lasciando invariata la riflettanza nello spettro del visibile. Questo evidenzia come la pressione selettiva degli impollinatori possa modellare le caratteristiche floreali.
Per quanto riguarda specifiche piante, si notano diverse complessità. Per esempio, la Binata dichotoma non è autoimpollinante, nonostante l'elevata frequenza con la quale l'evento si evidenzia porti molti neofiti a catalogarla erroneamente come tale. Il discorso impollinazione è complicato, al di là del vettore del polline e della fioritura contemporanea, c'è da tener conto della vitalità del polline, della sua scarsità o della sua sterilità. In alcune pratiche agricole, si consiglia l'uso di boro per aiutare la fecondazione nei fruttiferi, tra cui l'ulivo, con notevoli vantaggi.
Un altro esempio di complessità si riscontra nell'asimina, una pianta in cui lo stesso fiore passa da una iniziale fase femminile a una maschile. In questo caso, per ottenere raccolti notevoli, è possibile ricorrere all'impollinazione manuale, giocando "alle api, o meglio alle mosche", e trasferendo il polline dai fiori a campana aperta (fase maschile) a quelli a campana chiusa (fase femminile) con un pennello sottile. Esistono anche cloni autofertili, come il 'Cido', che semplificano la coltivazione, ma la presenza di diverse varietà vicine può favorire l'impollinazione incrociata anche per piante non autofertili.
Quando si lavora per scopi di selezione o ibridazione, può essere necessario sterilizzare i fiori. Per sterilizzare un tipo di fiore, si procede prima all'impollinazione dello stigma, poi, con una forbicina affilata, si recidono tutti gli stami con le antere. Per un altro tipo di fiore, dove gli stami sono posizionati all'interno rispetto agli stigmi, dopo aver impollinato, si sfili il cono e si recidono stami e antere. Questo permette un controllo preciso sulla genetica delle nuove piante.