Vi siete mai chiesti come si riproducono i pesci? La riproduzione e la gestazione nei pesci rappresentano uno degli aspetti più affascinanti della biologia marina. Diversamente dai mammiferi, i pesci presentano una varietà sorprendente di metodi di riproduzione, alcuni dei quali possono sembrare veramente alieni agli occhi umani. In questo articolo, esploreremo le strategie riproduttive uniche adottate dai pesci, mettendo in luce come queste tecniche contribuiscano alla loro sopravvivenza e proliferazione nelle acque di tutto il mondo. Nel meccanismo di nascita dei pesci, la fecondazione esterna è la modalità più diffusa, ma la storia evolutiva ha visto anche l'emergere di tecniche più complesse.
L'Evoluzione del Sesso e la Fecondazione Interna Primordiale
L'invenzione del sesso così come lo conosciamo sarebbe avvenuta 385 milioni di anni fa nei laghi fossili di Scozia, Estonia e Cina. I primi a sperimentare un rapporto sessuale tramite penetrazione e non attraverso fecondazione esterna, furono piccoli pesci ossei della classe dei Placodermi, pesci ormai estinti considerati gli antenati di molti vertebrati moderni. Questi erano antenati dotati di strutture riproduttive sorprendentemente avanzate per l'epoca. Protagonisti dello studio pubblicato su Nature sono i Microbrachius dicki, creature acquatiche lunghe 8 centimetri, vissuti nel periodo Devoniano per 70 milioni di anni, prima di estinguersi per motivi ancora ignoti.
Questi antichi pesci mostravano una piacevole novità evolutiva. In pratica, possedevano il precursore dei genitali maschili, utilizzato durante l'atto copulatorio per trasferire lo sperma nel corpo della femmina. I fossili femminili mostrano, di contro, una struttura ossea che doveva agire come "velcro", bloccando e tenendo in posizione l'organo maschile. La distribuzione filogenetica dei genitali maschili nella storia evolutiva dei vertebrati trova qui una delle sue prime espressioni. Maschi e femmine di Microbrachius si accoppiavano lateralmente, tenendosi in posizione con speciali pinne laterali. I pesci si trovano all’origine dell’albero genealogico dei placodermi: questo farebbe pensare che tutti i placodermi si riproducessero in questo modo. Studi precedenti avevano dimostrato che dobbiamo ai placodermi il nostro apparato masticatore e in ultima analisi anche il nostro sorriso.
The first ever recording of Grey Nurse Sharks/Raggies/Sand Tigers mating in the wild! Wolf Rock
Tuttavia, le cose non andarono sempre così. Molte specie ossee successive, discendenti dei placodermi, tornarono infatti alla fecondazione esterna, considerata più "primitiva" di quella interna. Questo rappresenta una sorta di passo indietro nella storia dell'evoluzione, prima d'ora ritenuto improbabile, ma che testimonia la straordinaria adattabilità dei meccanismi riproduttivi dei pesci.

Modalità Principali di Riproduzione nei Pesci Attuali
Cominciamo a parlare di come si riproducono i pesci suddividendo la riproduzione nelle tre macro-categorie principali: ovipara, vivipara e ovovivipara. A queste si aggiunge la riproduzione ermafrodita, che introduce un ulteriore livello di complessità biologica. La maturità sessuale dei pesci può avere durate molto diverse, con alcune specie che si riproducono anche dopo i dieci anni di età, e i cicli riproduttivi si ripetono per ogni stagione riproduttiva.
La Riproduzione Ovipara: La Prevalenza della Fecondazione Esterna
Fra le diverse modalità di riproduzione dei pesci, quella ovipara è sicuramente la più comune. In questo processo, i pesci depongono le loro uova all’esterno del proprio corpo, le quali verranno poi fecondate dal liquido spermatico rilasciato in acqua dal maschio. Parliamo dunque di fecondazione esterna: immediatamente dopo che le uova sono state deposte dalla femmina, il maschio rilascia il suo liquido spermatico nell’acqua, accanto ad esse. La fusione del nucleo dell'uovo con quello dello sperma segna l'inizio dello sviluppo. In seguito, dopo la fecondazione, le uova iniziano il loro processo di sviluppo. Questa modalità di riproduzione è tipica, ad esempio, del pesce rosso. Gli Osteitti hanno sessi quasi sempre separati e generalmente sono ovipari.
Vi sono sfumature diverse rispetto a come la riproduzione ovipara possa avvenire. Ad esempio, la femmina può deporre le uova liberamente nell’acqua, oppure farle aderire a piante, roccie o appositi nidi precedentemente preparati. Le uova possono essere deposte in una schiuma protettiva, in un nido, tra le alghe o tra coralli, o su altri substrati specifici. Alcune specie, come il pesce pagliaccio, depongono le uova su superfici solide e le sorvegliano attentamente; altre, come le anguille, depongono le uova in ambienti lontani, lasciandole al loro destino. Le uova possono essere adesive, galleggianti o affondanti, a seconda dell’habitat e della specie. Queste ultime due strategie si trovano spesso in ambienti instabili, dove la sopravvivenza dei piccoli richiede una maggiore protezione. La produzione di grandi quantità di gameti fa fronte all'alto tasso di mortalità e offre alcuni vantaggi, ad esempio la possibilità per le uova di essere rilasciate a gruppi un po' per volta nel corso del periodo riproduttivo. I tempi di schiusa variano secondo le dimensioni dell'uovo e della temperatura ambientale.
La Riproduzione Vivipara: Nascita di Piccoli Già Formati
Una seconda modalità di riproduzione è poi quella vivipara, in cui i pesci, anziché deporre le uova, partoriscono i loro piccoli completamente sviluppati. In questa modalità, gli embrioni si sviluppano all’interno del corpo della madre e ricevono nutrimento e ossigeno direttamente da essa attraverso una connessione ombelicale o altre strutture specializzate. Questi pesci, dunque, hanno una fecondazione interna, paragonabile a quella dei mammiferi. In contrasto, la fecondazione interna comporta l’inseminazione diretta delle uova all’interno del corpo femminile. Questo metodo è tipico di squali, razze e alcuni pesci ossei, come i guppy.

La riproduzione vivipara può variare da specie a specie. Alcuni pesci vivipari producono pochi piccoli alla volta, mentre altri possono dare alla luce un gran numero di avannotti (pesci piccoli). In alcune specie, i piccoli possono essere in grado di nuotare e nutrirsi autonomamente appena nati, mentre altre specie potrebbero richiedere ancora cure materne per un breve periodo di tempo. Questi pesci tendono a produrre un numero inferiore di uova, ma con tassi di sopravvivenza più elevati, grazie alla protezione offerta dal corpo materno o da altri meccanismi di salvaguardia. Nei pesci con fecondazione interna, le uova possono svilupparsi all’interno del corpo femminile, dove ricevono nutrimento diretto fino alla schiusa, ad esempio senza aderire all'utero, finché non giunge a completo sviluppo per essere espulso. Tale nutrimento può derivare anche da morti o tessuti materni in alcune specie. Nei Rajidae la riproduzione, dopo un corteggiamento rituale, comincia con la copula. Lo sperma viene conservato negli ovidotti in attesa delle uova da fecondare che discenderanno dall'ovaio. Subito dopo, la femmina secerne un rivestimento protettivo, per opera della ghiandola nidamentale, detta anche “ghiandola del guscio”. Nelle specie ovipare questo assume l'aspetto di una teca cornea molto resistente, mentre in quelle vivipare può ridursi a una membrana sottile, ma talvolta molto ampia, come si è scoperto nello squalo Carcharhinus dussumieri. In questa specie, infatti, le uova fecondate, del diametro di 20 mm, sono avvolte da una membrana che può superare i 40 cm di lunghezza e che viene mantenuta ripiegata nella parte anteriore dell’utero. Da qui, lentamente, questo “pseudo guscio” si svolge man mano che l’embrione aumenta di dimensioni.
In genere, dopo le fecondazioni, le fasi che portano alla formazione e allo sviluppo dell’embrione avvengono con regolarità. Questo, tuttavia, non impedisce ad alcune specie come lo squalo australiano dal muso affilato di bloccarne la crescita per un periodo di tempo che può protrarsi anche per sette mesi, allo scopo di far coincidere il parto con le condizioni ambientali più favorevoli ai nuovi nati.
La Riproduzione Ovovivipara: Un Compromesso Evolutivo
Vi è poi la riproduzione ovovivipara: un processo di riproduzione che combina caratteristiche sia della riproduzione ovipara che di quella vivipara. In questa modalità, gli embrioni si sviluppano all’interno del corpo della femmina, ma, a differenza della riproduzione vivipara, non ricevono nutrimento diretto dalla madre. Dopo la fecondazione, gli ovociti (ovuli) fecondati rimangono all’interno del corpo della madre e gli embrioni si sviluppano nutrendosi delle riserve presenti in questi ultimi. Queste riserve possono includere sostanze nutritive come il tuorlo, che forniscono energia e nutrienti necessari per lo sviluppo embrionale.

Nella riproduzione ovovivipara, non vi è, dunque, una connessione ombelicale tra la madre e gli embrioni. Gli embrioni si sviluppano indipendentemente, nutrendosi delle riserve presenti negli ovociti. La madre svolge un ruolo di protezione e sostegno, fornendo un ambiente favorevole allo sviluppo degli embrioni. Una volta completato lo sviluppo degli embrioni, la femmina partorisce i piccoli pesci completamente formati. I piccoli nascono vivi e sono in grado di nuotare e nutrirsi autonomamente fin dall’inizio. A differenza della riproduzione vivipara, non vi è alcun periodo di gestazione dopo il parto. La durata specifica della gestazione o incubazione è specifica per ogni specie, e va da 1 a 15 mesi.
La Riproduzione Ermafrodita: Flessibilità Biologica
Sapevate che alcuni pesci sono ermafroditi? Concludiamo poi con questo altro tipo di riproduzione. Nel caso di questo processo di riproduzione, un singolo individuo è in grado di fungere da maschio e da femmina, producendo sia gameti maschili che femminili. Esistono due tipi principali di riproduzione ermafrodita:
- Ermafroditismo simultaneo: Gli individui ermafroditi simultanei sono in grado di produrre e rilasciare sia gli ovociti che gli spermatozoi allo stesso tempo. Possono accoppiarsi con un altro individuo ermafrodita o con individui che seguono altri metodi di riproduzione. Questo tipo di riproduzione ermafrodita è comune in alcune specie di pesci tropicali come il pesce pagliaccio e il pesce chirurgo.
- Ermafroditismo sequenziale: Gli individui ermafroditi sequenziali cambiano il loro sesso durante il loro ciclo di vita. Nel caso del pesce pagliaccio, ad esempio, tutti gli individui nascono maschi e possono poi cambiare sesso.

L'Apparato Riproduttore dei Pesci: Organizzazione e Morfologia
La riproduzione di una specie ittica dipende sostanzialmente dalla funzionalità del suo apparato riproduttore. Questo è costituito da: gonadi che producono i gameti (maschili e femminili) e ormoni che condizionano il comportamento e lo sviluppo degli organi sessuali accessori e dei caratteri sessuali secondari; gonodotti per il trasporto di gameti o dello zigote; può comprendere, inoltre, alcuni derivati della cloaca o un poro genitale e organi genitali esterni implicati nell’accoppiamento. Il presente lavoro intende fornire una descrizione generale dell’organizzazione e della struttura delle gonadi nei pesci nell’ambito del contesto generale dello sviluppo in associazione alla descrizione dei processi di differenziazione sessuale di ovogenesi e spermatogenesi.
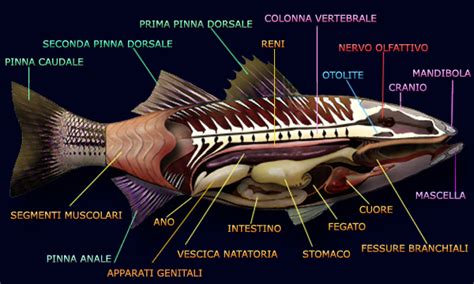
Le gonadi dei vertebrati sono composte da due tipi fondamentali di cellule: le cellule germinali, che danno luogo alla formazione di gameti maturi (uova e spermatozoi), e le cellule non germinali o somatiche, che danno sostegno, nutrimento e aiutano la regolazione dell’attività e lo sviluppo delle cellule germinali. Nelle gonadi sono presenti anche elementi di sostegno cellulari come le membrane basali. Nei Teleostei adulti, a parte alcune eccezioni, le gonadi sono strutture pari, allungate, attaccate alla cavità corporea da entrambi i lati del mesentere dorsale. La differenziazione sessuale è molto evidente, con variazioni somatiche spesso nelle dimensioni del corpo dei maschi, ma a volte anche delle femmine, più accentuate.
Le Gonadi Femminili: L'Ovaio e lo Sviluppo degli Ovociti
Le ovaie sono organi pari, di tipo sacciforme, ed occupano, una per parte, nella cavità celomatica, una posizione del tutto analoga a quella dei testicoli. Le strutture ovariche variano da relativamente semplici a organismi complessi. Racchiuse da una tunica albuginea contenente tessuto fibroso e muscolare liscio, appaiono usualmente come un piccolo grappolo di sfere bianco-arancioni nei pesci immaturi. Quando sono mature possono rappresentare fino al 70% del peso corporeo in alcune specie, mentre ad esempio nella trota iridea possono rappresentare circa il 15-18% del peso corporeo, e nella carpa circa il 32%. Ogni ovaio è appeso alla volta addominale per mezzo di un mesentere (mesovario) che inferiormente si rivolge verso l’alto a formare la piega genitale, la quale delimita la doccia paraovarica.
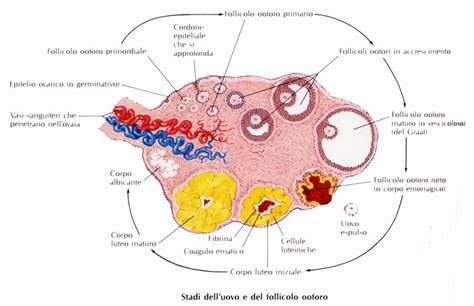
Nelle specie più progredite un breve condotto conduce le uova all’esterno attraverso l’apertura genitale. Le uova mature, infatti, cadono nella cavità celomatica e vengono convogliate in un canale formato dalla fusione della piega genitale sulle pareti del corpo detto ovidotto. Da qui, tramite energiche contrazioni dei muscoli della parete addominale, le uova vengono emesse all’esterno attraverso il poro genitale. Le specie più primitive (ad esempio, Salmonidi) non hanno un ovidotto completo e le uova vengono rilasciate in una piega del mesentere che eventualmente le rilascia poi in cavità peritoneale. Successivamente, raggiungono l’esterno attraverso l’apertura genitale. L’ovaio dei Teleostei è del tipo cistovarico (Hoar, 1969) cioè contiene un lume “ovocele” nel quale sono rilasciate le uova durante l’ovulazione. Pieghe ovigere, lamelle ovariche, contenenti i follicoli in via di sviluppo frequentemente si estendono nell’ovocele. Questo è un meccanismo contrario a quanto avviene nei vertebrati, nei quali cioè le uova vengono rilasciate al di fuori della gonade nella cavità corporea. La funzione degli ovidotti è quella di condurre le uova o le larve, nelle specie vivipare, al poro genitale. Tuttavia, nella femmina adulta di alcune specie come i Salmonidi, gli ovidotti regrediscono e le uova cadono all’interno della cavità prima di raggiungere il polo genitale, attraverso le rimanenze di un gonodotto a forma di imbuto. Prima dell’ovulazione le ovaie delle femmine mature sono degli organi relativamente grandi, occupanti la maggior parte della cavità corporea.
Microscopia Ovarica e il Follicolo Ovarico
Dal punto di vista morfologico funzionale l’unità basale dell’ovaio è il follicolo ovarico. L’organizzazione del follicolo ovarico è simile in tutti i Teleostei. L’ovocita localizzato centralmente è circondato da un involucro cellulare vitellino, conosciuto anche come zona radiata o corion. Questo involucro è a sua volta circondato da cellule follicolari che sono organizzate in un monostrato interno di cellule della granulosa e in uno strato esterno di cellule tecali. La membrana basale separa questi due strati cellulari.
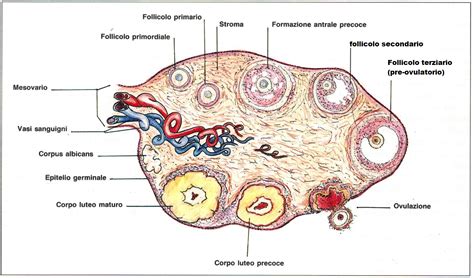
Negli ovari dei Teleostei possono essere ritrovati follicoli in tutti gli stadi di sviluppo, con momenti riproduttivi continui e prolungati (ad esempio, nel Syngnathus scovelli, il cavalluccio marino). Tuttavia, l’ovaio del cavalluccio marino non ha lamelle ovariche; esso è un tubo cilindrico delimitato da una parete ovarica esterna e un epitelio laminare interno. I follicoli ovarici si trovano tra questi due strati ma la loro distribuzione non è casuale. In sezione trasversale dell’ovaio i follicoli sono organizzati secondo uno stadio evolutivo. I follicoli più giovani originano dalla cresta germinale nella parte dorsale della gonade e follicoli progressivamente più vecchi si riscontrano in maniera ordinata attorno alla periferia. Follicoli pienamente maturi sono riscontrabili nella parte cosiddetta matura (cioè deputata alla maturazione ovocitica e all’ovulazione). Il parenchima dell’ovaio consiste di un tessuto stromale connettivale ben vascolarizzato contenente l’epitelio germinale e follicolare.
Nel tessuto follicolare producente ormoni, le cellule ovariche primitive o primarie rivestono cavità vuote o cavità potenziali, formate da pareti ripiegate in maniera complessa in cui vengono rilasciati gli oogoni che durante il processo ricevono uno strato di piccole cellule epiteliali: sono le cellule della granulosa, responsabili della formazione del tuorlo dell’uovo. Questa è la struttura del follicolo ovarico. Mano a mano che questo si accresce si forma tra l’ovocita e le cellule epiteliali una zona che si ispessisce gradualmente: la zona pellucida. Se un ovocita degenera prima dell’ovulazione, inizialmente viene invaso dalle cellule della granulosa e, successivamente, dai macrofagi.
Le cellule dello strato della granulosa risultano composte da una popolazione cellulare omogenea. Questo strato cellulare contiene anche le cellule micropilari, coinvolte nella formazione del micropilo. Il micropilo è un’apertura nell’involucro vitellino delle uova ovulate che permette l’accesso dello spermatozoo nella superficie dell’uovo durante la fertilizzazione. Lo strato delle cellule tecali è uno strato più eterogeneo composto di capillari, fibroblasti e spesso cellule tecali speciali. Delle proiezioni (microvilli) dall’ovocita e dalle cellule della granulosa penetrano l’involucro vitellino attraverso i canali del poro. Questi microvilli, specialmente quelli originati dall’ovocita, possono essere visti formare dei contatti con apposing cell type. In molte specie che sono state esaminate si formano delle giunzioni tipo gap eterocellulari nei punti di contatto tra microvilli di origine ovocitica e quelli di provenienza dalle cellule della granulosa. Le giunzioni gap sono strutture della membrana cellulare specializzate che danno continuità citoplasmatica tra le cellule. La loro funzione potrebbe essere quella di permettere lo scambio intercellulare diretto dei nutrienti e dei fattori che controllano lo sviluppo e la maturazione. Sono state descritte anche giunzioni tipo gap e tight omocellulari tra le cellule della teca, così come tra le cellule della granulosa nei follicoli ovarici dei Telostei.
L'Uovo di Pesce: Dimensioni, Struttura e Sviluppo
Le uova dei pesci ossei sono telolecitiche, ossia il vitello o tuorlo è molto sviluppato rispetto al citoplasma cellulare o macchia germinativa, e spesso presenta piccole gocce di grasso. La grandezza e il numero delle uova prodotte varia da specie a specie: da 1 mm nello Scoliodon a 100 mm negli squali nutrice (Ginglimostomatidi) e nello squalo dal collare Chlamydoselachus anguineus. Le uova possono essere prodotte in entrambe le ovaie come avviene nello squalo nello Squalus acanthias oppure soltanto in quello di destra, che è quella effettivamente funzionale nella maggior parte delle specie.

Nella trota iridea misurano circa 4-5,5 mm di diametro e sono in media 2.000 per ogni chilo di peso vivo; il loro diametro è proporzionale alla grandezza del riproduttore, ma il loro numero per chilo di peso vivo è più alto nei riproduttori di piccola mole (purché non al di sotto dei 200 grammi). Nella carpa hanno un diametro di 1,5-2 mm e sono deposte in numero di 200.000 per ogni chilo di peso vivo. Negli Elasmobranchi le uova rilasciate nella cavità addominale vengono sospinte dall’azione di cellule ciliate verso uno o entrambi gli ovidotti, lungo i quali discendono sino a quando non raggiungono la ghiandola nidamentale dove avviene la fecondazione e inizia la gestazione, oppure avviene la deposizione. In genere, dopo le fecondazioni, le fasi che portano alla formazione e allo sviluppo dell’embrione avvengono con regolarità. La temperatura ha un'elevata capacità di sopravvivenza per le uova. Una volta schiuso, il tuorlo presente nella sacca vitellina viene assorbito rapidamente. Successivamente, una serie di cambiamenti fa assumere alla larva l'aspetto dell'adulto (avannotto). Gli avannotti possono, però, essere molto diversi dall'adulto, come nel caso delle leptocefale delle anguille, che subiscono un'organogenesi e altre variazioni somatiche significative.
Le Gonadi Maschili: Il Testicolo e la Spermatogenesi
I testicoli appaiono come due formazioni allungate, di colore bianco latteo, che decorrono una per lato nella cavità celomatica, estendendosi dall’occipite fino all’apertura anale. Essi risultano sospesi tramite un mesentere (mesorchio) alla parete addominale dorsale adiacente alla vescica natatoria, se presente, e terminano assottigliati nei dotti spermatici che sboccano nel poro genitale. Nel maschio della trota iridea in fregola rappresentano circa il 3,33-3,9% del peso corporeo totale. La loro grandezza varia in relazione all’età del pesce e alla stagione: da piccole strisce nei giovani, possono diventare molto grandi negli adulti e arrivare fino al 12% del peso corporeo totale.
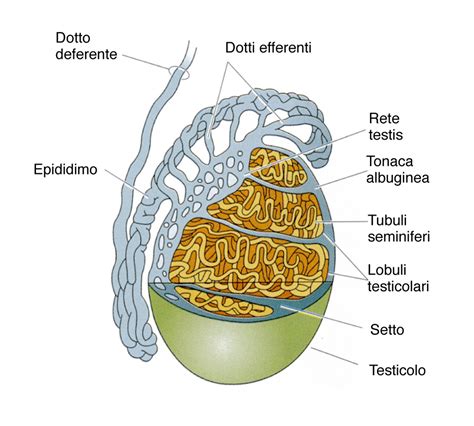
I testicoli si trovano all’interno di una sottile tunica albuginea e sono composti da una serie di tubuli seminiferi o sacchi a fondo cieco rivestiti con un epitelio spermatogenico, sia per tutta la loro lunghezza (come ad esempio nei Salmonidi e i Ciprinidi) o solamente nella parte terminale distale. La porzione caudale dei testicoli di specie come gli Ictaluridi e alcuni Ciprinidi è rivestita da cellule epiteliali secretorie non germinali, che possono contribuire allo stoccaggio e nutrizione degli spermatozoi e alla formazione dello sperma. L’epitelio spermatogenico produce gli spermatociti che eventualmente possono andare incontro a divisione meiotica e produrre gli spermatozoi. Gli spermatozoi, che si differenziano in testa e in coda e sono dotati di movimento, vengono nutriti dalle cellule del Sertoli fino al loro rilascio. Nella maggior parte delle specie gli spermatozoi sono emessi da ciascun testicolo attraverso il dotto deferente. I due condotti si uniscono caudalmente e gli spermatozoi vengono rilasciati nell’ambiente attraverso l’apertura genitale a livello di papilla urinaria. Questi condotti sono assenti in alcune specie relativamente primitive come i salmonidi, dove gli spermatozoi vengono rilasciati nella cavità corporea prima di emetterli all’esterno attraverso l’apertura genitale.
Classificazione e Struttura Microscopica del Testicolo
L’organizzazione dei testicoli è diversa da quella dell’ovaio e riflette una situazione anatomica e funzionale diversa nello sviluppo dei gameti maschili. In passato i testicoli dei Teleostei sono stati classificati come appartenenti ai tipi spermatogenici ristretto o non ristretto, basati cioè sulla localizzazione degli spermatozoi (Grier, 1981) o ai tipi giugulare o tubulare in base alla presenza o assenza del lumen (Billard, 1986). In queste classificazioni i tipi spermatogenici ristretto o non ristretto corrispondono rispettivamente ai tipi testicolari giugulare e tubulare.
Più recentemente Callard (1991), utilizzando un approccio filogenetico più ampio, ha classificato i testicoli di tutti i vertebrati come tubulari (Mammiferi, Uccelli e Rettili) o loculari (Teleostei, Anfibi). Nella classificazione di Callard un tubulo è un compartimento germinale aperto dei testicoli, mentre un loculo è un sacco a fondo cieco. Così molti testicoli dei Teleostei vengono ad essere classificati di tipo loculare, malgrado differenze nel modello della distribuzione spermatogonica o la presenza o assenza di un lume all’interno del loculo. Grier nel 1993 si è trovato in accordo con la classificazione di Callard (1991), ma ha descritto un nuovo tipo di testicolo tubulare riscontrato nei Teleostei: il testicolo tubulare anastomizzante. Testicoli tubulari anastomizzanti sono riscontrati in Teleostei filogeneticamente meno evoluti (ad esempio, Lepisosteus platyrhincus, Ictalurus atalis, Opisthonema oglinum, Dorosoma petense, Esox niger, ecc…) e sono caratterizzati da una rete ramificata di tubuli che, come i tubuli testicolari dei Mammiferi, formano delle anse alla periferia della gonade (Grier, 1993). Nel presente lavoro è stata adottata la classificazione di Callard del 1991 come modificata da Grier nel 1993 in testicoli lugulari e tubulari anastomizzanti. Comunque, l’enfasi sarà maggiormente sui testicoli lugulari, poiché sono a disposizione relativamente poche informazioni sui testicoli lugulari anastomizzanti.
I loguli testicolari nei Teleostei sono demarcati da una membrana basale e da uno strato sovrastante di cellule confinanti (Mioidi). I loguli contengono gli spermatocisti che sono unità sferiche composte di cellule germinali e cellule del Sertoli. Gli spermatocisti sono delimitati parzialmente o completamente a seconda della specie da una membrana basale. Le cellule germinali all’interno di una spermatocisti sono collegate citoplasmaticamente da ponti intercellulari e la loro differenziazione è abbastanza sincrona. Nelle specie con loguli vuoti (come la maggior parte dei Teleostei) gli spermatogoni così come gli spermatocisti a vari stadi di sviluppo, possono essere visti attraverso la maggior parte della lunghezza del logulo e gli spermatozoi maturi sono rilasciati al momento della produzione spermatica all’interno del lume logulare. Tuttavia, nei testicoli con loguli solidi pieni, come ad esempio negli Atheriniformi, l'organizzazione può differire.
Cura Parentale e Sopravvivenza della Prole
Il processo di sviluppo delle uova può richiedere diversi tipi di attenzione o protezione. Alcuni pesci, ad esempio, proteggono le proprie uova all’interno della bocca o delle camere branchiali, mentre altri pesci non offrono alcun tipo di cure parentali. Molti pesci mostrano notevoli livelli di cura parentale. Ciò può includere la protezione del nido, dove molti pesci difendono aggressivamente le uova da predatori e parassiti. Alcuni genitori puliscono regolarmente le uova per prevenire la formazione di funghi e batteri. In alcune specie, i genitori trasportano le uova in bocca per proteggerle fino alla schiusa, un fenomeno chiamato incubazione orale. Alcuni sorvegliano il nido, altri proteggono le uova tra le pinne o addirittura nella bocca.

Nel caso degli ippocampi (o cavallucci marini), i ruoli si invertono: è il maschio a rimanere “incinto”, portando le uova in una sacca specializzata. La sopravvivenza delle giovani generazioni, la prole nei primi giorni di vita, è fondamentale per il mantenimento delle popolazioni ittiche. Strategie come la produzione massiva di uova e la cura parentale specializzata aiutano a garantire che una percentuale sufficiente di larve raggiunga l’età adulta. Una maggiore produzione di fitoplancton e zooplancton, ad esempio, è vantaggiosa per i nuovi nati e la loro crescita.
Fattori Ambientali e Impatto Umano sulla Riproduzione
I cambiamenti ambientali stanno mettendo a rischio la riproduzione dei pesci. Anche la pesca eccessiva, l’uso di reti a strascico e la distruzione delle barriere coralline stanno riducendo le zone di riproduzione. Questi fattori influenzano direttamente la capacità delle specie ittiche di perpetuarsi, alterando gli habitat naturali e le risorse alimentari disponibili per la prole. La perdita di substrati idonei per la deposizione delle uova o la nidificazione rappresenta una minaccia significativa.

tags: #pesci #ossei #fecondazione #esterna