Nell'ultimo decennio, l'interesse per la biologia della riproduzione si è notevolmente accresciuto, in parte grazie ai successi ottenuti nell'applicazione delle colture gametiche ed embrionali alle pratiche mediche, veterinarie e biotecnologiche, e in parte a causa delle pressanti esigenze della società attuale. Nella scienza medica, le tecnologie della riproduzione assistita (Assisted Reproductive Technologies, ART) sono state messe a punto soprattutto per porre rimedio alla sterilità, mentre nella zootecnia il crescente fabbisogno alimentare di una popolazione mondiale in piena espansione ha stimolato la ricerca di metodi per incrementare la produzione e l'efficienza riproduttiva del bestiame. La più antica applicazione documentata di una tecnica di riproduzione assistita risale al 1783, quando il fisiologo italiano Lazzaro Spallanzani inseminò artificialmente una cagna ottenendo una cucciolata; ma fu solo intorno al 1900 che il russo I. I. Ivanov e la sua scuola misero a punto vagine artificiali e tecniche di inseminazione per equini, bovini e ovini. La grande potenzialità dell'inseminazione artificiale negli animali da allevamento dipende dal fatto che l'eiaculato maschile contiene diversi milioni di spermatozoi, sufficienti, in teoria, a fecondare centinaia di femmine. Un decisivo passo avanti in questa direzione fu compiuto verso la fine degli anni quaranta, quando, a Cambridge (Inghilterra), il gruppo di ricercatori diretto da Chris Polge sviluppò tecniche di congelamento e di conservazione di spermatozoi animali. Quasi contemporaneamente furono messi a punto i metodi per isolare e manipolare i gameti femminili: il primo esperimento di maturazione in vitro degli oociti risale infatti a oltre mezzo secolo fa, allorché si osservò che l'oocita primario del coniglio riattiva spontaneamente la meiosi quando viene liberato dal suo follicolo e posto in un idoneo mezzo di coltura. Il primo bambino ‛concepito' in vitro è nato nel 1978; attualmente gli individui IVF (In Vitro Fertilized) sono circa 200.000 in tutto il mondo.
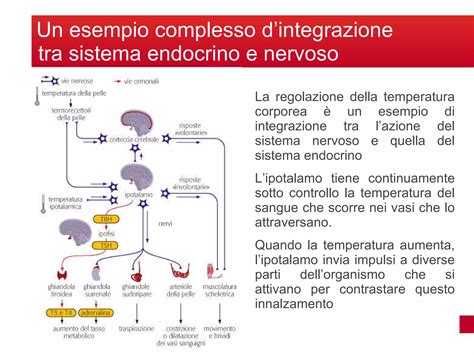
Il Controllo Endocrino della Riproduzione
Il fatto che, nel maschio, l'eccitamento sessuale, l'erezione e l'eiaculazione siano fenomeni posti sotto il controllo cerebrale risulta ovvio; quel che appare meno ovvio è che anche i cicli ovarico e testicolare siano coordinati a livello centrale. Per molti anni dopo la scoperta degli ormoni gonadotropi ipofisari - l'ormone follicolo-stimolante (Follicle Stimulating Hormone, FSH) e l'ormone luteinizzante (Luteinizing Hormone, LH) -, l'ipofisi anteriore, o adenoipofisi, fu considerata un organo autonomo, finché esperimenti su animali in cui erano state indotte lesioni nell'ipotalamo dimostrarono in modo chiaro come il sistema nervoso mediasse i processi riproduttivi. L'ipotalamo, una piccola parte del cervello limitata anteriormente dalla lamina terminale telencefalica e posteriormente dal mesencefalo, controlla i cicli sessuali, la crescita, la gravidanza, l'allattamento e varie altre reazioni basali ed emotive. Nonostante le sue piccole dimensioni, l'ipotalamo è una struttura estremamente complessa. Ogni funzione è associata a una o più aree dell'ipotalamo, costituite da aggregazioni di neuroni denominate ‛nuclei ipotalamici'. Nel contesto della riproduzione, vari gruppi di nuclei ipotalamici sono collegati alla sottostante ghiandola pituitaria (o ipofisi) mediante connessioni neurali e vascolari.
L'ormone liberante l'ormone gonadotropo (Gonadotrophin Releasing Hormone, GnRH) è il prodotto neurosecretorio di neuroni ipotalamici e viene trasportato all'adenoipofisi attraverso vasi portali. L'ormone GnRH, un decapeptide con struttura piro-Glu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly-NH2, è il principale mediatore della riproduzione da parte del sistema nervoso centrale, e ogni difetto nella sua sintesi, nel suo immagazzinamento, nel suo rilascio o nella sua azione indurrà un'inibizione parziale o completa della funzione gonadica. Esso viene secreto in modo pulsante e si lega ai recettori specifici presenti sulla membrana plasmatica delle cellule basofile ipofisarie secernenti gonadotropine (o cellule gonadotrope) per attivare il sistema di trasduzione del segnale, che usa l'inositolo-trifosfato come secondo messaggero intracellulare. Tale segnale induce il movimento dei granuli secretori verso la membrana plasmatica e infine la secrezione pulsante di LH e FSH. L'esposizione continua a GnRH o a un suo analogo produce l'occupazione stabile del sito di legame del recettore, il disaccoppiamento del recettore stesso dal suo sistema di trasduzione del segnale e infine la riduzione della secrezione di LH e FSH.
Nelle femmine dei Primati, la produzione di gonadotropine è regolata dall'ovario. Modesti livelli circolatori di estradiolo esercitano un controllo a retroazione negativa sulla produzione di LH e di FSH, mentre, se presente a livelli stabilmente elevati, l'estradiolo esercita un controllo retroattivo positivo. Alti livelli plasmatici di progesterone amplificano gli effetti retroattivi negativi dell'estradiolo e mantengono la secrezione di FSH e di LH a un livello modesto. La secrezione di FSH, ma non di LH, è regolata anche da molecole non steroidee, proteiche, ad alto peso molecolare (circa 30.000), denominate ‛inibine' e rinvenute nel liquido follicolare.
I meccanismi neuroendocrini che regolano la funzione testicolare sono fondamentalmente simili a quelli che regolano l'attività ovarica. L'unità ipotalamo-ipofisaria nel maschio è responsabile della secrezione di gonadotropine che modulano le attività endocrina e spermatogenica del testicolo e sono esse stesse soggette a regolazione retroattiva. Una delle maggiori differenze tra il maschio e la femmina consiste nel fatto che il processo di produzione dei gameti e degli ormoni steroidei, dopo la pubertà, nel maschio è continuo, mentre nella femmina è ciclico. Un riflesso diretto di ciò è l'assenza di regolazione retroattiva positiva dei prodotti testicolari sul rilascio di gonadotropina. Nel maschio, LH stimola la produzione, da parte delle cellule di Leydig, di testosterone che regola a sua volta la secrezione di LH, riducendone la frequenza e l'ampiezza dei picchi. Sebbene meno chiaramente che nella femmina, anche nel maschio (in estratti testicolari) sono state rinvenute molecole simili all'inibina, che probabilmente svolgono un ruolo nella regolazione dell'FSH.
La Produzione dei Gameti
Per comprendere i problemi associati alla fecondazione è necessario prima esaminare come vengano prodotti i gameti. Nel maschio che ha raggiunto la pubertà, le cellule germinali - fino ad allora bloccate in interfase - iniziano a proliferare per mitosi, seguita dalla meiosi e da una graduale riorganizzazione dei componenti cellulari, caratterizzata da perdita di citoplasma. Si è calcolato che nel mammifero adulto vengono prodotti circa cinquecento spermatozoi al secondo per ogni grammo di testicolo. Le cellule staminali, o spermatogoni A0, si trovano nella parte basale dell'epitelio germinativo del tubulo seminifero. Da questa popolazione si sviluppano periodicamente spermatogoni A1 che vanno incontro a un numero fisso di divisioni mitotiche per formare un clone di cellule figlie. Esistono prove sperimentali a favore dell'ipotesi che uno dei cloni di cellule figlie possa fungere da centro secondario di espansione clonale (focus) di cellule staminali. Dopo l'ultima divisione mitotica hanno origine gli spermatociti primari che si spostano verso il compartimento luminale dell'epitelio ed entrano in meiosi; in seguito a due divisioni meiotiche si formano prima due spermatociti secondari figli e infine quattro spermatidi. Sebbene i nuclei degli spermatidi contengano serie aploidi di cromosomi, gli autosomi continuano a sintetizzare modeste quantità di RNA messaggero, RNA ribosomiale e proteine. Il DNA degli spermatidi diviene a questo punto altamente condensato ed è infine strettamente associato a protammine. La riorganizzazione citoplasmatica dà origine alla coda, alla parte centrale che contiene i mitocondri, all'acrosoma e al corpo residuo, in cui è confinato il citoplasma in eccesso; il modellamento dello spermatozoo è probabilmente regolato dalle cellule del Sertoli.
Nei Mammiferi, gli spermatozoi che lasciano il testicolo non sono in grado di fecondare gli oociti, ma acquistano questa capacità passando attraverso l'epididimo, in un processo denominato ‛maturazione epididimale'. Gli spermatozoi testicolari, anche se opportunamente trattati e posti in soluzione fisiologica, sono essenzialmente privi di motilità. La capacità di movimento è probabilmente regolata a livello della membrana plasmatica, poiché la sua rimozione e l'esposizione ad ATP, cAMP e Mg2+ induce la motilità. Si ritiene che il trasferimento dal liquido epididimale di carnitina e di una proteina responsabile del movimento in avanti sia importante per lo sviluppo della motilità dello spermatozoo. Poiché l'osmolalità e la composizione chimica del liquido epididimale variano da un segmento al successivo lungo l'epididimo, è probabile che la membrana plasmatica dello spermatozoo sia modificata passo dopo passo mentre percorre il condotto. La testa dello spermatozoo acquisisce quindi la capacità di aderire alla zona pellucida dell'oocita con un incremento della carica negativa netta. Durante la maturazione, lo spermatozoo consuma le riserve endogene di substrati metabolici, diventando dipendente dalle risorse esogene, come il fruttosio, e perde la goccia citoplasmatica. Quando lascia il testicolo lo spermatozoo è ricoperto da numerose macromolecole che, durante il passaggio attraverso l'epididimo, vengono perdute o modificate, mentre se ne aggiungono di nuove. Le più importanti fra queste molecole sono le glicoproteine, sintetizzate grazie all'azione di enzimi quali la galattotransferasi e la sialtransferasi. Le modificazioni della capacità di legare lectine, che la membrana plasmatica dello spermatozoo subisce durante la maturazione epididimale, sono un segnale delle alterazioni del residuo terminale saccaridico di queste glicoproteine.
Nella femmina, la fase di proliferazione mitotica termina prima della nascita e tutti gli oogoni entrano nella prima divisione meiotica, diventando oociti primari. Alla pubertà sono disponibili per l'attività riproduttiva circa 200.000 cellule germinali. Durante la prima profase meiotica gli oociti sono circondati da cellule mesenchimali ovariche a costituire i follicoli primordiali. Gli oociti si bloccano allo stadio di diplotene, con il grande nucleo caratteristico, chiamato ‛vescicola germinale'. Non è noto come questi oociti quiescenti restino vitali per una cinquantina d'anni (fino alla menopausa). Lo sviluppo di alcuni di questi follicoli primordiali ha luogo per la prima volta nella pubertà: il follicolo cresce da venti a parecchie centinaia di micron e l'oocita in esso contenuto da dieci a circa cento micron; la fase di accrescimento coinvolge essenzialmente la sintesi e l'immagazzinamento di grandi quantità di RNA, proteine e substrati metabolici. Durante la crescita, le circostanti cellule della membrana granulosa vanno incontro a divisione mitotica e nella regione compresa tra l'oocita e queste cellule viene secreto un rivestimento glicoproteico: la zona pellucida. Giunzioni comunicanti (gap junctions) tra l'oocita e le proiezioni citoplasmatiche delle cellule accessorie che penetrano nella zona pellucida permettono il trasferimento dei substrati e delle informazioni necessarie allo sviluppo.
I pochi follicoli che hanno completato la loro fase di crescita sono chiamati ‛follicoli precavitari' o primari; molti vanno incontro ad atresia. Le gonadotropine circolanti inducono la conversione dei follicoli precavitari in follicoli cavitari o di Graaf. La fase precavitaria dura, nella donna, 8-12 giorni, durante i quali nel follicolo inizia ad accumularsi un liquido chiaro, il liquor folliculi, in cui è sospeso l'oocita con le circostanti cellule del cumulo ooforo. Durante questa fase di crescita il follicolo produce una quantità crescente di androgeni e di estrogeni. Una successiva ondata di LH induce un veloce ulteriore accumulo di liquido, il quale nella successiva fase preovulatoria, della durata di circa 36 ore, farà raggiungere al follicolo, che verrà poi ovulato, le sue dimensioni definitive, pari, nella donna, a 25 mm di diametro. Si ritiene che le fimbrie dell'ovidotto espellano quindi l'oocita nell'ampolla, dove avverrà l'ovulazione. In quest'ultimo stadio dell'ovogenesi la membrana nucleare si dissolve, la meiosi viene reiniziata e il primo globulo polare viene espulso: la cellula femminile è ora nella seconda metafase. Contemporaneamente ha inizio un processo di maturazione citoplasmatica, che comprende un decremento nella conduttanza di potassio, la depolarizzazione della membrana plasmatica e la migrazione dei granuli corticali verso la superficie dell'oocita.
LA FECONDAZIONE UMANA
Coito e Fecondazione
Nei Mammiferi la fecondazione è interna e il gamete maschile deve quindi essere introdotto nel tratto genitale femminile durante il coito, che varia, nella durata, da minuti, nella specie umana, a ore, nei cammelli, ma è sempre accompagnato da molte modificazioni fisiologiche. L'erezione del pene nella specie umana può essere indotta da stimoli tattili e psichici, per esempio visivi. Il fenomeno erettile si basa sulla diminuzione della resistenza e la conseguente dilatazione delle arterie profonde del pene nonché sulla chiusura delle comunicazioni arterovenose e delle valvole venose. In condizioni di vasocongestione, i testicoli possono aumentare di volume del 50%. L'eiaculazione del liquido seminale è provocata dalla contrazione dei muscoli lisci dell'uretra e dei muscoli striati del pene; tale contrazione è ritmica e induce il mescolamento del liquido prostatico, ricco di fosfatasi acida, con la frazione del vaso deferente contenente gli spermatozoi e la frazione della vescichetta seminale contenente il fruttosio.
Nella donna, la stimolazione tattile del glande clitorideo e della parete vaginale porta alla congestione della vagina e delle grandi labbra e a un incremento delle dimensioni vaginali. Nell'uomo, nel coniglio, nella pecora, nel bue e nel gatto il seme viene eiaculato nella vagina; nel maiale, nel cane e nel cavallo è invece deposto direttamente nella cervice uterina e nell'utero stesso. In molte specie il seme coagula rapidamente dopo la deposizione nel tratto genitale femminile, in conseguenza dell'interazione con un enzima prostatico. La coagulazione potrebbe servire per trattenere gli spermatozoi nella vagina o per proteggerli dal suo ambiente acido (pH µ 5,7). Entro pochi minuti dall'accoppiamento, nella cervice dell'utero si possono rinvenire spermatozoi, ma, nella donna, il 99% di essi non raggiunge l'utero.
Il miglioramento genetico e la riproduzione degli animali d’allevamento rappresenta un aspetto cruciale per il mantenimento degli animali e la sostenibilità economica delle aziende coinvolte. Le biotecnologie della riproduzione, a partire dall’inseminazione artificiale fino alle biotecnologie basate sull’utilizzo degli embrioni, hanno avuto e hanno tutt’ora un grosso impatto nell’allevamento animale su scala mondiale e soprattutto nei paesi sviluppati. Lo sfruttamento della genetica femminile per la selezione e il miglioramento genetico passa attraverso le tecnologie dell’embrione. In passato la tecnica più utilizzata era stata quella basata sull’induzione farmacologica di ovulazioni multiple (superovulazione) e la raccolta degli embrioni tramite lavaggio uterino. Le tecnologie in vitro stanno sostituendo per i notevoli vantaggi che offrono la superovulazione, e il numero di embrioni prodotti con tecnologie in vitro nella specie bovina sta rapidamente crescendo in tutto il mondo, soprattutto per i vantaggi offerti dalle nuove tecniche di selezione basate sulla genomica. Anche in altre specie quali, cavallo e piccoli ruminanti, l’ovum pick up e la produzione in vitro degli embrioni trovano applicazioni crescenti come pure nella specie suina l’impiego di ovociti maturi per biotecnologie come la clonazione o l’ingegneria genetica. Tuttavia in queste specie meno studiate la tecnologia in vitro non dà gli stessi risultati che si ottengono nel bovino ed esiste quindi un ampio margine di miglioramento. L’ovocita rappresenta il “componente biologico” che maggiormente incide nel determinare il successo dello sviluppo embrionale con la nascita di animali.
L'Inseminazione Artificiale
L'inseminazione artificiale è una pratica molto diffusa negli allevamenti. In un mercato ad alta richiesta di animali, come quello della carne, delle uova e del latte, e in un mercato altamente redditizio come quello dell’ippica e, in parte, dell’allevamento cinotecnico, questa pratica apporta numerosi benefici in termini economici. Nell’allevamento di cani con pedigree, la pratica è diffusa ma non d’obbligo, mentre negli altri settori è quasi ormai l’unico metodo di fecondazione degli animali. È estremamente raro, se non nei pochi casi “isole felici”, che per gli animali da produzione o nell’ippica professionale venga praticata la monta naturale. Nella società odierna, concedere un accoppiamento naturale tra gli animali d’allevamento, ad esempio, dove la produzione raggiunge le migliaia di unità l’anno e dove il bilancio costi/introiti è calcolato all’ultimo centesimo, sarebbe completamente impensabile. Da un punto di vista etico appare come una barbarità e uno snaturamento degli animali; fermiamoci però a pensare che sono le meccaniche intrinseche dei mercati, e quindi l’alta domanda di animali, e indirettamente le abitudini dei consumatori, che alimentano questa pratica.
La fecondazione artificiale è la metodica che permette di depositare gli spermatozoi nell’apparato riproduttore femminile nel momento più propizio, a seconda della specie animale, e di instaurare una gravidanza. Ad oggi interessa il settore zootecnico (bovini, suini, ovicaprini, ad esempio), dell’ippica ed in parte il mondo cinotecnico, ancor più frequentemente nel caso di alcune razze come i brachicefali di piccola taglia (bull dog francese, bull dog inglese, eccetera). La pratica della fecondazione artificiale si struttura in quattro fasi: prelievo del seme (del toro, stallone, ariete, cane, eccetera) attraverso l’utilizzo di vagine artificiali; controllo della qualità del seme in termini di numero di spermatozoi, loro mobilità, ad esempio, e sua diluizione con prodotti specifici; conservazione del seme diluito tramite congelamento o refrigerazione; scongelamento (nel caso) e inseminazione attraverso il deposito del seme nell’apparato riproduttivo della femmina. Tutto il procedimento, in ogni sua fase, avviene tramite materiali e tecnologie specifiche ed ormai altamente specializzate per aumentarne le performance e ridurre al minimo il margine d’errore (fecondazione non avvenuta con successo) e la possibilità di provocare lesioni agli animali, si pensi ad esempio alle mucose dell’apparato riproduttore femminile nel momento dell’inseminazione. Parallelamente alle tecnologie per la fecondazione artificiale, si sono sviluppate e perfezionate le tecnologie per ottimizzare la rilevazione dell’estro delle femmine, per poter ottimizzare la buona riuscita della pratica.
Perché si ricorre a questa pratica? Tra queste le principali sono:
- Promuovere la genetica di qualità: La selezione dei riproduttori maschi e la loro valutazione genotipica consentono agli allevatori di ottenere rese più alte. Esempi sono un’alta conversione in carne nel caso dei bovini da carne, una buona genetica nella filiera del latte, campioni di gare nei casi delle corse di cavalli o elevati standard morfologici di razza nel mondo cinofilo.
- Aumentare l’efficienza del riproduttore: Si utilizzano pochi esemplari maschi, ad alta qualità di prestazione per l’ambito di appartenenza, per un numero elevato di femmine di una stessa specie, o razza. Fecondare con il seme diluito di alta qualità di un unico animale diventa vantaggioso nel mercato sia in termini economici che quantitativi, perché si può raggiungere un numero di figli molto elevato, maggiore rispetto alla monta naturale.
- Consentire risultati di fecondazione molto elevati rispetto alla monta naturale: Essendo una pratica molto accurata.
- Prevenire le lesioni che si verificano durante la monta naturale, sia per i maschi che per le femmine. Come ad esempio scivolamenti, contusioni, morsi, ed altre lesioni.
- Evitare la diffusione delle infezioni a trasmissione sessuale, data dall’assenza di contatto tra le mucose degli animali ed il controllo sanitario del seme.
L'Inseminazione in Vitro
L'inseminazione in vitro (IVF) rappresenta un'evoluzione significativa rispetto all'inseminazione artificiale tradizionale. Sebbene il concetto di base rimanga la fecondazione assistita, l'IVF sposta il processo dal corpo dell'animale al laboratorio. Questo approccio offre una serie di vantaggi, tra cui un maggiore controllo sulle condizioni di fecondazione e la possibilità di selezionare gli embrioni più vitali. Il processo inizia con la raccolta degli oociti maturi dalla femmina, spesso attraverso procedure minimamente invasive come l'ovum pick-up (OPU). Questi oociti vengono poi messi in coltura in specifici terreni che mimano l'ambiente fisiologico del tratto riproduttivo femminile. Successivamente, gli spermatozoi vengono preparati e aggiunti agli oociti in una piastra di coltura. La fecondazione avviene quando uno spermatozoo penetra nell'oocita. Gli embrioni risultanti vengono quindi coltivati in laboratorio per alcuni giorni, permettendo loro di svilupparsi fino a uno stadio precoce (spesso blastocisti). Gli embrioni più promettenti vengono poi trasferiti nell'utero della femmina ricevente, con l'obiettivo di ottenere una gravidanza.
L'IVF ha trovato applicazioni estese in diverse specie, in particolare nei bovini, dove ha contribuito significativamente al miglioramento genetico e alla produzione di animali di alta qualità. Tuttavia, il suo successo varia a seconda della specie, con sfide ancora presenti in specie meno studiate come cavalli, piccoli ruminanti e suini. Nonostante ciò, il campo è in continua evoluzione, con nuove tecniche e protocolli che vengono sviluppati per ottimizzare i tassi di successo. L'importanza dell'oocita come "componente biologico" chiave nel determinare il successo dello sviluppo embrionale e la nascita di animali è ampiamente riconosciuta, sottolineando la necessità di una gestione attenta e di tecniche precise nella sua manipolazione.
LA FECONDAZIONE UMANA
La Micromanipolazione
La micromanipolazione rappresenta un insieme di tecniche avanzate che permettono di manipolare gameti ed embrioni a livello microscopico. Queste procedure sono fondamentali per alcune delle applicazioni più sofisticate della riproduzione assistita, come l'iniezione intracitoplasmatica dello spermatozoo (ICSI) e la biopsia embrionale. L'ICSI, in particolare, prevede l'iniezione di un singolo spermatozoo direttamente all'interno dell'oocita. Questa tecnica è estremamente utile in casi di grave infertilità maschile, dove la motilità o il numero di spermatozoi sono insufficienti per una fecondazione in vitro convenzionale. La micromanipolazione consente anche la biopsia embrionale, che prevede la rimozione di una o poche cellule da un embrione in uno stadio precoce di sviluppo. Queste cellule possono poi essere utilizzate per analisi genetiche pre-impianto (PGT), permettendo di identificare eventuali anomalie cromosomiche o mutazioni genetiche prima del trasferimento dell'embrione nell'utero. Questo approccio è particolarmente importante nella medicina umana per ridurre il rischio di trasmettere malattie genetiche ereditarie, ma trova applicazione anche in zootecnia per la selezione di animali geneticamente superiori o esenti da determinate patologie.
Le Tecnologie Future
Il futuro della riproduzione nei mammiferi si prospetta ricco di innovazioni. La ricerca continua a esplorare nuove frontiere, con l'obiettivo di migliorare ulteriormente l'efficienza, la precisione e l'accessibilità delle tecnologie riproduttive. Tra le aree di maggiore interesse vi sono lo sviluppo di nuove tecniche di crioconservazione dei gameti e degli embrioni, che potrebbero permettere una conservazione più a lungo termine e con minori danni cellulari. Si stanno studiando anche metodi per migliorare la maturazione in vitro degli oociti e la coltura embrionale, al fine di aumentare i tassi di successo delle procedure IVF. Un'altra area promettente è la genetica riproduttiva, che include tecniche come la clonazione e l'ingegneria genetica, che potrebbero permettere la creazione di animali con caratteristiche desiderate. Inoltre, l'applicazione di intelligenza artificiale e machine learning all'analisi dei dati riproduttivi potrebbe portare a diagnosi più accurate e a piani di trattamento personalizzati. L'obiettivo finale è quello di ottimizzare la riproduzione in tutte le specie, sia per scopi conservazionistici che per migliorare la produzione animale e la salute umana.
"Avantea S.r.l. In “termini utilitaristici”, gli animali sono fonte di compagnia per alcuni, fonte di alimentazione per molti, fonte di svago per altri, ed arricchiscono, completano ed accompagnano le nostre vite. Troppo spesso ne sono però vittime, basti pensare alla cattività, agli allevamenti intensivi, alla spinta genetica cui li sottoponiamo, allo svago senza senso come le corse di cani o la tauromachia. Un esempio è la fecondazione artificiale, specialmente nell’ambito degli allevamenti intensivi e nell’ambito dell’ippica. Pochi, davvero pochi, di noi si saranno soffermati a pensare come avviene la riproduzione tra gli animali che vivono sotto il controllo umano, per i quali sono richiesti numeri enormi di nascituri o altissimi standard performativi."
tags: #inseminazione #nei #mammiferi