Lo sviluppo della vita, dalla sua origine più microscopica fino alla formazione di un organismo complesso e funzionale, rappresenta uno dei fenomeni più affascinanti e intricati in biologia. Il processo mediante il quale l'embrione si forma e si sviluppa è definito sviluppo embrionale, o embriogenesi. Questa incredibile sequenza di eventi trasforma una singola cellula in una miriade di altre, organizzate in tessuti, organi e sistemi, che culminano nella nascita di un nuovo individuo. Sebbene le strategie riproduttive e gli ambienti in cui si sviluppano siano estremamente diversi tra le varie classi di vertebrati, quali pesci, anfibi, rettili, uccelli e mammiferi, i principi fondamentali dell'embriogenesi mostrano sorprendenti somiglianze, pur con affascinanti adattamenti specifici.
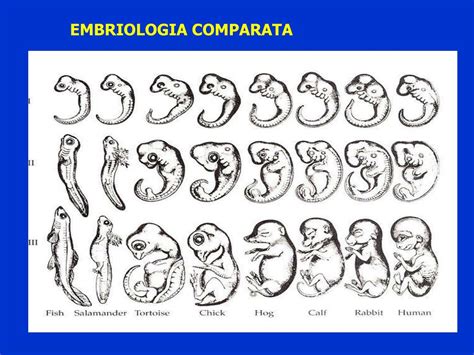
Questo viaggio nell'embriogenesi comincia sempre dalla fecondazione, l'unione di due cellule sessuali, e procede attraverso stadi ben definiti di divisione cellulare, organizzazione tissutale e differenziazione degli organi, per culminare nella formazione di un organismo completo, pronto per la vita al di fuori dell'ambiente embrionale. Ogni passaggio è finemente regolato da meccanismi genetici e molecolari che, pur variando nei dettagli, seguono schemi evolutivi conservati, testimoniando la profonda interconnessione della vita sulla Terra.
La Fecondazione: Il Primo Passo dello Sviluppo
Lo sviluppo embrionale comincia con la fecondazione della cellula uovo aploide, da parte di uno spermatozoo anch'esso aploide. Questo evento critico, che segna l'inizio di una nuova vita, implica l'incontro e la fusione di due gameti, ciascuno portatore di metà del patrimonio genetico necessario alla specie. Nei vertebrati, la fecondazione può avvenire con modalità diverse, distinguendosi principalmente in interna ed esterna.
Nei pesci e negli anfibi, la fecondazione è spesso esterna. Questo significa che le uova vengono deposte nell'ambiente acquatico, e gli spermatozoi vengono rilasciati nelle vicinanze per fecondarle. Questo processo richiede una sincronizzazione precisa tra i partner e può essere influenzato da fattori ambientali. Nonostante la vastità dell'ambiente acquatico, gli spermatozoi devono comunque compiere un viaggio verso l'uovo, guidati da segnali chimici e meccanici.
Al contrario, nei rettili, negli uccelli e nei mammiferi, la fecondazione è interna. Gli spermatozoi vengono depositati all'interno del tratto riproduttivo femminile, fornendo un ambiente più protetto e controllato per l'incontro con l'uovo. Anche in questi casi, il percorso dello spermatozoo verso l'uovo è irto di ostacoli e richiede un'efficienza notevole. Ad esempio, dei circa 300 milioni di spermatozoi umani eiaculati durante il coito, solo 200 circa raggiungono l'ovidotto. Questa drastica riduzione del numero sottolinea la selettività del processo e la necessità di superare barriere fisiologiche significative.
Una volta raggiunto l'ovidotto o le immediate vicinanze dell'uovo, gli spermatozoi devono superare alcuni strati di cellule granulose che circondano la cellula uovo. Questa barriera cellulare è sostenuta da una matrice extracellulare complessa. La presenza dell'enzima ialuronidasi legato alla membrana cellulare anteriore dello spermatozoo, lo aiuta ad attraversare la matrice extracellulare che circonda le cellule granulose. L'azione dell'enzima digerisce l'acido ialuronico che compone la matrice extracellulare e gli permette di aggirare queste cellule.
Superato questo primo strato, gli spermatozoi devono attaccarsi e attraversare la zona pellucida, uno strato glicoproteico acellulare che avvolge la membrana plasmatica dell'uovo. Nei pressi della cellula uovo, lo spermatozoo si attacca alla zona pellucida attraverso un complesso meccanismo che coinvolge numerose proteine di membrana specifiche. Questo legame induce una serie di eventi cruciali nello spermatozoo, nota come reazione acrosomiale. L'attivazione di queste proteine induce una cascata di eventi intracellulari nello spermatozoo, che si concludono con l'esocitosi della vescicola acrosomica. Il contenuto enzimatico della vescicola, ricco di enzimi proteolitici, si riversa dunque verso la zona pellucida e inizia a digerirla, creando un passaggio. Questo permette allo spermatozoo di far passare lo spermatozoo verso la cellula uovo e favorirne la fusione cellulare.
I meccanismi molecolari responsabili del processo di fusione delle due membrane plasmatiche, quella dello spermatozoo e quella dell'uovo, non sono del tutto chiari, sebbene si sia scoperta l'esistenza di due proteine necessarie al processo: Izumo1 e CD9. La fusione delle membrane attiva la cellula uovo, un evento fondamentale che previene ulteriori fecondazioni. Questa attivazione si manifesta con un rapido aumento della concentrazione di calcio nel citosol della cellula uovo. Quest'aumento di cationi è responsabile della depolarizzazione della membrana plasmatica dell'uovo, un cambiamento elettrico che agisce come un blocco rapido alla polispermia, allontanando efficacemente gli altri spermatozoi. Tutto ciò serve a prevenire eventi di polispermia (più fecondazioni da parte di diversi spermatozoi), che sarebbero letali per lo sviluppo dell'embrione, poiché porterebbero ad un eccesso di materiale genetico.
La fecondazione si conclude con la fusione del materiale genetico dello spermatozoo con quello della cellula uovo. Questo evento finale unisce i due pronuclei aploidi, riformando un nucleo diploide. La nuova cellula che si origina prende il nome di zigote ed è una cellula diploide che possiede tutte le informazioni genetiche necessarie a sviluppare un intero organismo funzionante. È il punto di partenza per ogni successivo stadio dello sviluppo embrionale, un momento di incredibile potenziale biologico.
Dalla Cellula Singola alla Pluricellularità: La Segmentazione
Una volta formatosi, lo zigote inizia un processo di rapida divisione cellulare noto come segmentazione. La morfologia dello zigote dipenderà principalmente dalla forma della cellula uovo, in quanto lo spermatozoo apporta a quest'ultima solo il materiale genetico contenuto nel suo nucleo e i centrioli. La dimensione della cellula uovo e, soprattutto, la quantità e la distribuzione del vitello (materiale nutritivo) all'interno di essa, sono fattori determinanti per la modalità di segmentazione.
Il processo che porta lo zigote alla pluricellularità è chiamato segmentazione. Questa fase consiste in una serie di divisioni mitotiche che si distinguono per un'assenza di accrescimento cellulare tra una divisione e l'altra. Tipicamente, le cellule attraversano cicli cellulari completi con fasi G1, S, G2 e M. Tuttavia, durante la segmentazione, queste sono conseguenti divisioni mitotiche senza accrescimento, ovvero senza le fasi G1 e G2. Ciò significa che il volume totale dell'embrione non aumenta in questa fase, ma viene semplicemente suddiviso in un numero crescente di cellule più piccole. Queste divisioni portano lo zigote a dividersi (segmentarsi) in 2 cellule figlie chiamate blastomeri, poi in 4, in 8, in 16 e così via, in una progressione esponenziale.
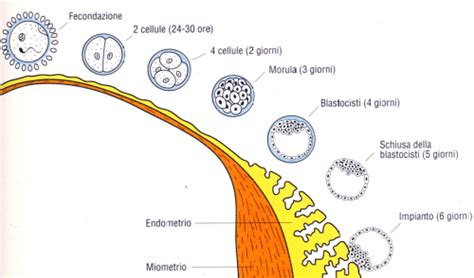
La modalità di segmentazione può variare significativamente tra le classi di vertebrati, in relazione alla quantità di vitello.
- Mammiferi: Le uova dei mammiferi sono tipicamente microlecitiche (poverissime di vitello). La segmentazione è oloblastica (divisione completa della cellula) e rotazionale, il che significa che il piano di divisione delle blastomere non è sempre parallelo o meridionale. Dopo che la settima divisione ha prodotto 128 cellule, l'embrione prende il nome di Blastula. Nei mammiferi in questa fase formano una struttura chiamata blastocisti, caratterizzata da una massa cellulare interna che è distinta dalla blastula circostante. Questa massa cellulare interna (ICM) darà origine all'embrione vero e proprio, mentre le cellule trofoblastiche circostanti formeranno le membrane extraembrionali e parte della placenta. Nel topo, le cellule germinali primordiali derivano da uno strato di cellule nella massa cellulare interna della blastocisti (l'epiblasto) come risultato di un'ampia riprogrammazione dell'intero genoma.
- Anfibi: Le uova degli anfibi sono mesolecitiche (con una quantità moderata di vitello, concentrato nel polo vegetativo). La segmentazione è anch'essa oloblastica ma è disuguale. Le divisioni sono più rapide e le blastomere più piccole nel polo animale (povero di vitello) rispetto al polo vegetativo (ricco di vitello). Questo porta alla formazione di una blastula con una cavità, il blastocele, spostata verso il polo animale.
- Pesci, Rettili e Uccelli: Le uova di queste classi sono macrolecitiche (ricchissime di vitello). A causa dell'abbondante vitello, la segmentazione è meroblastica o parziale, il che significa che solo una piccola porzione citoplasmatica nel polo animale (il blastodisco) si divide. Il resto dell'uovo, composto quasi interamente dal vitello, rimane indiviso. Questo tipo di segmentazione è chiamato discoidale, poiché le divisioni avvengono solo sul disco citoplasmatico. Anche in questi casi, si forma una struttura che può essere considerata una blastula, ma è un disco di cellule appoggiato sul vitello indiviso. Ad esempio, l'uovo di uccello, con il suo enorme vitello, è un classico esempio di segmentazione meroblastica discoidale.
Queste differenze nella segmentazione riflettono l'adattamento delle specie alle diverse strategie riproduttive e alle risorse nutritive disponibili all'embrione in via di sviluppo. Indipendentemente dalla modalità, l'obiettivo finale della segmentazione è produrre un sufficiente numero di cellule, i blastomeri, che si auto-organizzano per formare la blastula o la blastocisti, la struttura che sarà la base per le successive, più complesse, riorganizzazioni cellulari.
L'Organizzazione dei Foglietti Embrionali: La Gastrulazione
Dopo la segmentazione, l'embrione subisce una serie di movimenti cellulari drammatici e riorganizzazioni che portano alla formazione dei foglietti embrionali. Il processo che porta la blastula ad essere un embrione triblastico, ossia dotato di 3 foglietti embrionali, è detto gastrulazione. Questi tre foglietti embrionali primari - ectoderma, mesoderma ed endoderma - sono le fondamenta da cui si differenzieranno tutti i tessuti e gli organi dell'organismo adulto.
La gastrulazione è un periodo di intensa attività morfogenetica, durante il quale le cellule della blastula si muovono e si riposizionano attivamente. Le modalità precise della gastrulazione variano notevolmente tra i gruppi di vertebrati, ancora una volta influenzate dalla quantità e dalla distribuzione del vitello nell'uovo.
- Anfibi: Negli anfibi, la gastrulazione avviene attraverso l'invaginazione delle cellule nel blastoporo, una depressione che si forma nella regione equatoriale dell'embrione. Le cellule che migrano all'interno formeranno il mesoderma e l'endoderma, mentre quelle che rimangono all'esterno costituiranno l'ectoderma. Questa invaginazione è un esempio classico di riorganizzazione tridimensionale.
- Uccelli e Rettili: In queste classi, data la segmentazione meroblastica e la formazione di un blastodisco, la gastrulazione è caratterizzata dalla formazione di una struttura chiamata stria primitiva. Questa stria, che si forma sulla superficie del blastodisco, agisce come un punto di ingresso attraverso il quale le cellule superficiali migrano verso l'interno, andando a formare il mesoderma e l'endoderma. L'ectoderma rimane lo strato più esterno. La stria primitiva è un omologo funzionale del blastoporo degli anfibi.
- Mammiferi: Anche nei mammiferi, la gastrulazione avviene attraverso la formazione di una stria primitiva, simile a quella di uccelli e rettili, sebbene avvenga all'interno della massa cellulare interna della blastocisti. Le cellule dell'epiblasto migrano attraverso la stria primitiva per formare i tre foglietti germinativi.
Indipendentemente dalla modalità specifica, il risultato finale della gastrulazione è un embrione organizzato in tre strati. L'ectoderma è il foglietto più esterno e darà origine all'epidermide, al sistema nervoso e ad altre strutture associate. L'endoderma è il foglietto più interno e formerà il rivestimento del tratto gastrointestinale e i suoi organi accessori, nonché il rivestimento delle vie respiratorie. Il mesoderma, il foglietto intermedio, darà origine a muscoli, scheletro, sistema circolatorio, reni e altri tessuti connettivi.
Durante la gastrulazione, o immediatamente dopo, la blastula si trasforma e comincia a delineare le prime strutture assiali del corpo. Specificamente, la superficie più esterna (ectoderma) si alza fino a formare un ponte, la cresta neurale, che chiudendosi va a formare il tubo neurale. Questo è il primo abbozzo che poi si svilupperà nella colonna vertebrale e nel midollo spinale che corre al suo interno. Questo processo, noto come neurulazione, è il primo passo verso la formazione del sistema nervoso centrale.
Contemporaneamente alla neurulazione, si delinea anche il primitivo sistema digestivo. La blastula invagina una porzione di ectoderma (che diventerà poi la bocca) fino a collegarla ad un'altra porzione invaginata (l'ano) e formando di fatto un tubo continuo che si svilupperà poi durante l'organogenesi nel tubo digerente. L'intero tubo digerente è ricoperto dall'endoderma, confermando la sua origine dal foglietto germinativo più interno. Questa precoce formazione di un asse corporeo e dei primi abbozzi di organi è un momento cruciale che stabilisce il piano corporeo di base dell'organismo.
la fecondazione e impianto
La Morfogenesi Assiale e la Formazione dei Somiti: La Somitogenesi
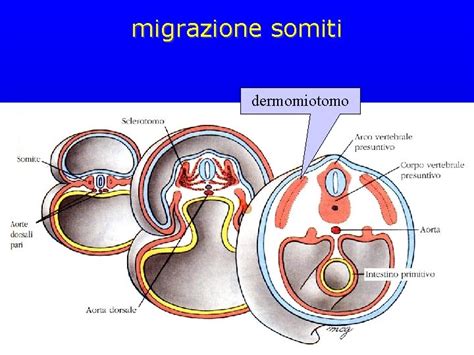
Con la definizione dei tre foglietti embrionali, l'embrione entra in una fase di maggiore complessità strutturale, caratterizzata dalla formazione di strutture segmentate lungo l'asse antero-posteriore. La somitogenesi è il processo mediante il quale vengono prodotti i somiti, che sono segmenti primitivi del mesoderma parassiale. Questi somiti sono blocchi di mesoderma che si formano simmetricamente su entrambi i lati del tubo neurale e della notocorda e sono fondamentali per lo sviluppo del piano corporeo segmentato di tutti i vertebrati.
La somitogenesi inizia con la formazione di somitomeri che segnano i futuri somiti nel mesoderma presomitico, una regione del mesoderma che non è ancora segmentata. Il mesoderma presomitico dà origine a coppie successive di somiti, di aspetto identico che si differenziano negli stessi tipi di cellule ma le strutture formate dalle cellule variano a seconda della parte antero-posteriore. Questo significa che, sebbene i somiti diano origine a tessuti simili (come muscoli, vertebre e derma), la loro posizione lungo l'asse del corpo determinerà la loro specifica differenziazione e le caratteristiche morfologiche che assumeranno. Ad esempio, le vertebre toraciche hanno costole, le vertebre lombari no; questa diversità è radicata nella differente identità posizionale dei somiti da cui derivano.
Verso la fine della seconda settimana dopo la fecondazione, nell'embrione umano, inizia la segmentazione trasversale del mesoderma parassiale, che viene convertito in una serie di masse ben definite, più o meno cubiche, note anche come somiti. Questi somiti occupano l'intera lunghezza del tronco su entrambi i lati della linea mediana dalla regione occipitale della testa fino alla coda. Il ritmo e il numero di somiti formati sono specie-specifici e finemente controllati da un "orologio di segmentazione" molecolare.
Ogni segmento, o somite, contiene una cavità centrale che, tuttavia, viene presto riempita di cellule angolari a forma di fuso. Questa riorganizzazione interna è l'inizio della differenziazione delle cellule dei somiti in tre principali compartimenti: lo sclerotomo, che darà origine alle vertebre e alle costole; il miotomo, che formerà i muscoli scheletrici; e il dermatomo, che contribuirà alla formazione del derma della pelle.
I somiti giacciono immediatamente sotto l'ectoderma sull'aspetto laterale del tubo neurale e della notocorda e sono collegati al mesoderma laterale dalla massa cellulare intermedia. La loro posizione strategica li rende cruciali per l'interazione con il tubo neurale e la notocorda, che sono fonti di segnali di sviluppo che guidano la differenziazione dei somiti stessi.
Il numero di somiti varia tra le diverse specie di vertebrati, riflettendo le differenze nella lunghezza e nella complessità della colonna vertebrale. Nell'uomo, quelli del tronco possono essere disposti nei seguenti gruppi, vale a dire: cervicale 8, toracico 12, lombare 5, sacrale 5 e coccigeo da 5 a 8. Quelli della regione occipitale della testa sono generalmente descritti come quattro. Similmente, nei rettili, uccelli, anfibi e pesci, i somiti si formano in modo sequenziale, stabilendo la segmentazione del corpo e fornendo i precursori per la muscolatura assiale e scheletrica, che sono essenziali per il movimento e il supporto strutturale dell'organismo. La loro formazione ordinata è un segno distintivo dell'embriogenesi dei vertebrati, garantendo una precisa organizzazione spaziale delle strutture corporee.
La Nascita degli Organi: L'Organogenesi e la Neurulazione
Dopo che i diversi strati germinali sono stati definiti con la gastrulazione e la segmentazione del mesoderma parassiale ha dato origine ai somiti, l'embrione entra nella fase di organogenesi. Ad un certo punto, dopo che i diversi strati germinali sono stati definiti, inizia l'organogenesi. Questa fase è caratterizzata dalla differenziazione e dalla formazione degli organi e dei sistemi corporei a partire dai tre foglietti embrionali. È un periodo di rapida crescita e modellamento, dove le cellule si specializzano e si organizzano in strutture complesse e funzionali.
Un aspetto fondamentale dell'organogenesi nei vertebrati è la formazione del sistema nervoso centrale. Il primo stadio nei vertebrati è chiamato neurulazione, dove la placca neurale, una regione ispessita dell'ectoderma dorsale, si piega su se stessa formando il tubo neurale. Questo processo è cruciale in tutte le classi di vertebrati, dai pesci ai mammiferi, e getta le basi per lo sviluppo del cervello e del midollo spinale. Anomalie nella neurulazione possono portare a gravi difetti congeniti del sistema nervoso.
Durante l'organogenesi, i tre foglietti germinativi interagiscono in modi complessi per guidare la formazione di ogni organo:
- Ectoderma: Oltre a formare il tubo neurale e quindi il sistema nervoso centrale, l'ectoderma darà origine all'epidermide della pelle, ai peli, alle unghie, alle ghiandole cutanee, all'epitelio sensoriale degli occhi, delle orecchie e del naso, e all'enamelo dei denti. Le cellule della cresta neurale, derivate dall'ectoderma nella regione dorsale del tubo neurale, sono estremamente versatili e migrano in tutto il corpo per formare una vasta gamma di tessuti e organi, inclusi i neuroni periferici, le cellule pigmentate della pelle, le cartilagini e le ossa della faccia e del cranio.
- Mesoderma: Questo strato si differenzia in una moltitudine di tessuti. Il mesoderma parassiale, dopo la somitogenesi, forma lo sclerotomo (scheletro assile), il miotomo (muscolatura scheletrica) e il dermatomo (derma). Il mesoderma intermedio forma i reni e le gonadi. Il mesoderma della lamina laterale si divide in uno strato somatico (che contribuirà alle pareti del corpo e agli arti) e uno strato splancnico (che formerà la muscolatura liscia del tratto digestivo e le membrane sierose degli organi interni, oltre al cuore e al sistema circolatorio).
- Endoderma: Questo foglietto interno forma l'epitelio di rivestimento del tratto gastrointestinale e respiratorio, nonché le ghiandole associate a questi sistemi, come il fegato, il pancreas e le ghiandole tiroide e paratiroide.
Le tempistiche e le specifiche di organogenesi variano notevolmente tra le classi di vertebrati, spesso influenzate dal loro ambiente di sviluppo.
- Pesci e Anfibi: Molti pesci e anfibi depongono uova che si sviluppano esternamente e spesso passano attraverso uno stadio larvale. Nella maggior parte degli animali l'organogenesi, insieme alla morfogenesi (la formazione della forma complessiva dell'organismo), si traduce in una larva. Queste larve (come i girini degli anfibi o gli avannotti dei pesci) sono spesso organismi autonomi che si nutrono e crescono, ma non assomigliano all'adulto e devono subire una metamorfosi per raggiungere la forma adulta. Questo stadio larvale è un adattamento che permette di sfruttare diverse nicchie ecologiche e risorse alimentari.
- Rettili e Uccelli: Questi animali si sviluppano in uova cleidoiche (con guscio) che contengono abbondante vitello e membrane extraembrionali protettive (amnios, corion, sacco vitellino, allantoide). Lo sviluppo è diretto, senza uno stadio larvale distinto. L'organogenesi porta direttamente alla formazione di un giovane che assomiglia a una versione in miniatura dell'adulto, sebbene possa ancora richiedere cure parentali significative. La presenza di un uovo con guscio ha permesso a queste specie di colonizzare ambienti terrestri.
- Mammiferi: Nei mammiferi placentati, l'embrione si sviluppa all'interno del corpo materno, nutrito dalla placenta. Anche qui lo sviluppo è diretto, senza stadio larvale. L'organogenesi è un processo continuo e altamente coordinato, che si conclude con la nascita di un individuo che, pur immaturo, è una versione completa e funzionante del genitore. La gestazione interna offre protezione e un ambiente stabile per l'organogenesi.
L'organogenesi è il culmine di tutti i processi embrionali precedenti, in cui le cellule si specializzano, si organizzano in tessuti e formano organi distinti. È un periodo critico in cui la corretta interazione tra i diversi tessuti e i segnali di sviluppo è essenziale per la formazione di un organismo sano e vitale. L'efficacia di questo processo determina la funzionalità dell'individuo e la sua capacità di sopravvivere e riprodursi nel proprio ambiente.
Interazioni Celulari e Molecolari nello Sviluppo Embrionale
Ogni fase dell'embriogenesi, dalla fecondazione all'organogenesi, è orchestrata da un complesso dialogo di segnali cellulari e molecolari. I processi descritti, sebbene possano sembrare una sequenza lineare di eventi, sono in realtà profondamente interconnessi e dipendenti l'uno dall'altro. La precisione con cui questi eventi si susseguono è il risultato di milioni di anni di evoluzione, che hanno selezionato meccanismi di sviluppo robusti e adattabili.
I meccanismi molecolari responsabili del processo di fusione delle due membrane plasmatiche all'inizio dello sviluppo, come l'interazione tra le proteine Izumo1 e CD9 durante la fecondazione, sono solo un esempio della specificità e della complessità che caratterizzano ogni singola fase. Questi meccanismi garantiscono che l'unione dei gameti sia selettiva e che l'uovo venga attivato in modo appropriato per prevenire la polispermia, un evento altrimenti letale.
Durante la segmentazione, il controllo del ciclo cellulare è drasticamente alterato. Le cellule si dividono rapidamente senza periodi di crescita (fasi G1 e G2 del ciclo cellulare) al fine di aumentare il numero di blastomeri senza aumentare le dimensioni complessive dell'embrione. Questo meccanismo di divisione rapida è essenziale per generare un numero sufficiente di cellule che possano poi riorganizzarsi durante la gastrulazione. La regolazione di questi cicli cellulari accelerati è controllata da proteine specifiche che vengono attivate e disattivate in momenti precisi.
La gastrulazione, con i suoi movimenti cellulari massivi e coordinati, è guidata da segnali di adesione e migrazione cellulare. Le cellule si muovono seguendo gradienti chimici e fisici, interagendo con la matrice extracellulare e con altre cellule. La formazione della stria primitiva nei mammiferi, uccelli e rettili, o il blastoporo negli anfibi, è il risultato di movimenti cellulari concertati che portano all'internalizzazione di alcune popolazioni cellulari e alla stratificazione dei foglietti germinativi. Questi processi sono finemente regolati da geni omeotici e da vie di segnalazione come Wnt, TGF-beta e Hedgehog, che stabiliscono l'identità e il destino delle cellule.
la fecondazione e impianto
La somitogenesi, con la sua caratteristica formazione segmentata, è un esempio di come i segnali oscillatori ("orologi di segmentazione") possano essere tradotti in schemi spaziali. La formazione sequenziale dei somiti è regolata da un'interazione tra il "clock" e il "wavefront" di espressione genica, garantendo che ogni somite si formi nel posto giusto e al momento giusto. L'identità di ciascun somite lungo l'asse antero-posteriore, che determina se darà origine a una vertebra cervicale, toracica o lombare, è stabilita dai geni Hox. Questi geni, altamente conservati tra le specie, codificano per fattori di trascrizione che "istruiscono" le cellule sul loro posizionamento lungo l'asse corporeo.
Infine, l'organogenesi è un processo di differenziazione cellulare e di interazione induttiva tra tessuti. Un tessuto induce un altro tessuto a differenziarsi in una specifica direzione. Ad esempio, la notocorda induce la placca neurale a piegarsi e a formare il tubo neurale. Questo dialogo continuo tra tessuti diversi è fondamentale per la formazione di strutture complesse come gli occhi, gli arti o il cuore. Le cellule devono non solo specializzarsi, ma anche auto-organizzarsi e connettersi in reti funzionali. I segnali tra l'ectoderma, il mesoderma e l'endoderma sono intricati e reciprocamente dipendenti, assicurando che gli organi si formino correttamente e siano posizionati in modo appropriato.
In tutte le specie di vertebrati, dal più semplice pesce all'uomo, queste interazioni molecolari e cellulari sono la forza trainante dietro la straordinaria trasformazione di una singola cellula fecondata in un organismo multicellulare altamente organizzato. Le variazioni osservate tra le classi riflettono gli adattamenti a specifici ambienti e strategie riproduttive, ma i principi sottostanti di regolazione genica, segnalazione cellulare e morfogenesi rimangono sorprendentemente conservati, evidenziando l'unità fondamentale della vita.
tags: #embrioni #animali #rettili #mammiferi #uccelli #anfibi