La vita complessa degli organismi eucarioti, inclusi gli esseri umani, è intrinsecamente legata a un processo cellulare straordinario e intricato noto come meiosi. Questo meccanismo di divisione cellulare è specificamente coinvolto nelle cellule germinali e riveste un'importanza capitale per la riproduzione sessuata e per la perpetuazione della diversità genetica. A differenza della mitosi, che genera cellule figlie geneticamente identiche alla cellula madre, la meiosi si distingue per il dimezzamento del corredo cromosomico da doppia copia a singola copia, producendo quattro cellule finali con un patrimonio genico diverso fra loro. Questa diversità è il risultato della combinazione casuale di cromosomi di provenienza materna e paterna, una caratteristica che riflette la generazione dei nonni del potenziale nuovo individuo.
Negli animali, ogni genitore contribuisce a questo processo fornendo, attraverso la meiosi, un corredo cromosomico in copia unica. Tale corredo si trova nel gamete femminile, l'ovulo (o ovocita), e nel gamete maschile, lo spermatozoo. La fusione di questi due corredi cromosomici durante la fecondazione ricostituisce un corredo completo, dando origine alla prima cellula di un nuovo individuo, lo zigote. Grazie alla ricombinazione genetica, ogni zigote si presenta ogni volta diverso dai precedenti e dai genitori, evidenziando il ruolo cruciale della meiosi nella variabilità di specie. La meiosi è, in sintesi, quel complicato processo di divisione, tipico degli organismi eucarioti, al termine del quale una cellula diploide diventa aploide, un passo essenziale per la formazione dei gameti.
Le Fasi della Meiosi I: La Divisione Riduzionale
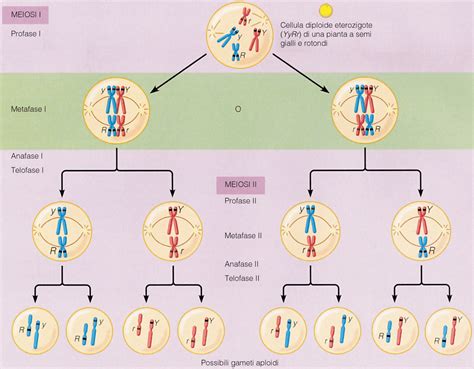
Prima dell'inizio della meiosi, avviene la duplicazione del materiale genetico. In questa fase preliminare, ogni cromosoma risulta formato da due cromatidi fratelli identici, uniti in corrispondenza del centromero mediante molecole di coesina. La prima divisione meiotica, o meiosi I, è chiamata riduzionale. Il suo scopo è generare, a partire da una cellula (2n), due cellule definibili aploidi in base al numero di cromosomi, i quali risultano però formati da due cromatidi.
La Profase I: Il Palcoscenico della Ricombinazione
La meiosi I si apre con la profase I, riconosciuta come il processo più lungo e complicato dell'intera divisione meiotica. Questa fase è suddivisa in diverse sottofasi, ognuna con eventi caratteristici che preparano la cellula alla segregazione cromosomica e alla ricombinazione genetica:
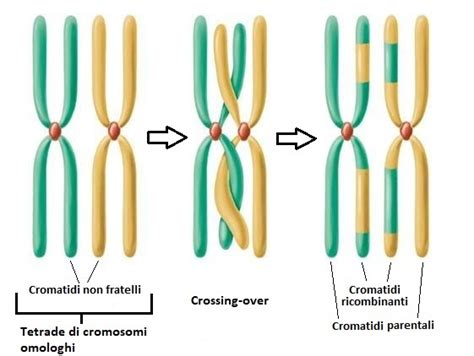
- Leptotene: In questa sottofase, il materiale genetico si condensa a formare strutture bastoncellari, visibili come filamenti sottili e allungati, che non sono ancora scissi longitudinalmente. Durante il leptotene avvengono i DSB (Double Strand Breaks), punti di rottura controllata della doppia elica del DNA. Questi punti corrispondono alle zone dove si potrà andare incontro al crossing-over nelle fasi successive, un evento chiave per la variabilità genetica.
- Zigotene: È lo stadio durante il quale avviene la sinapsi, il processo mediante il quale i cromosomi omologhi si mettono in coppia. Questo appaiamento forma una struttura denominata bivalente (o tetrade o duplex), in cui ciascun cromosoma è formato da due cromatidi, e quindi quattro cromatidi formano una tetrade.
- Pachitene: Si distingue in "precoce", dove si completa l'appaiamento degli omologhi, e "avanzato", in cui i cromosomi si accorciano, si inspessiscono. In questa fase avviene il crossing-over, un processo mediante il quale i cromatidi non fratelli di cromosomi omologhi si scambiano parti equivalenti, determinando nuove combinazioni di geni e favorendo l'evoluzione. Tuttavia, il crossing-over non è ancora visibile, poiché i cromosomi rimangono in stretto contatto fra loro.
- Diplotene: In questo stadio, i cromosomi omologhi di ciascun bivalente cominciano a separarsi (desinapsi), soprattutto a livello del centromero, per la progressiva scomparsa del complesso sinaptinemale. Nonostante la separazione, i due cromosomi di ciascuna coppia di omologhi restano in contatto grazie a connessioni chiamate chiasmi. Questi chiasmi sono i segni visibili dell'avvenuto crossing-over e hanno un importante ruolo strutturale. Negli umani, in condizioni normali, in ogni coppia di omologhi deve avvenire per forza almeno un chiasma. Il risultato visibile del crossing-over è, infatti, una struttura a croce chiamata chiasma.
- Diacinesi: Nel corso di questa sottofase, i cromosomi completano la loro condensazione e diventano chiaramente visibili.
Durante la profase I, si sviluppa anche il fuso meiotico, una struttura cruciale per la segregazione cromosomica. Il fuso è costituito da due coppie di centrioli, situate ai poli opposti della cellula, da cui fuoriescono fibre di microtubuli. Tali fibre agganciano i cromosomi mediante il cinetocore, una piastra proteica situata a livello del centromero. Le fibre del fuso si collegano ai cromosomi: ogni cromosoma, diviso in 2 cromatidi tenuti insieme dal centromero, è legato tramite gli asteridi alle fibre del fuso.
Metafase I, Anafase I e Telofase I
Dopo la complessa profase I, i bivalenti si allineano sulla piastra metafasica durante la Metafase I. Successivamente, in Anafase I, i cromosomi omologhi si separano e migrano verso i poli opposti della cellula. È importante notare che, a differenza della mitosi, in questa fase non si separano i cromatidi fratelli, ma interi cromosomi omologhi. La Telofase I può variare a seconda della specie. Ai poli opposti della cellula si cominciano a formare i nuclei e avviene la citodieresi, con la conseguente scissione cellulare. I microtubuli del fuso scompaiono. Il risultato sono due cellule aploidi, ma con ogni cromosoma ancora costituito da due cromatidi fratelli.
La Meiosi II: La Divisione Equazionale
La seconda divisione meiotica, o meiosi II, ha un carattere equazionale, molto simile a una normale divisione mitotica. Essa ha il compito di separare i cromatidi fratelli all'interno di ciascuna delle due cellule figlie prodotte dalla meiosi I.
Profase II, Metafase II, Anafase II e Telofase II
La Profase II è una fase breve, durante la quale i cromosomi, ancora costituiti da due cromatidi, si ricondensano e ricompare il fuso meiotico. Compaiono nuovamente le fibre del fuso che agganciano i cinetocori dei cromosomi. Nella Metafase II, i cromosomi si allineano sulla piastra metafasica in ciascuna delle due cellule. In Anafase II, i cromatidi fratelli si separano, diventando cromosomi indipendenti, e migrano verso i poli opposti delle cellule. Infine, nella Telofase II, si formano i nuovi nuclei attorno ai cromosomi che hanno raggiunto i poli, e la citodieresi divide ciascuna delle due cellule in due, portando alla formazione di un totale di quattro cellule aploidi, ognuna con un corredo cromosomico singolo e non duplicato. La telofase I può variare a seconda della specie. Compaiono nuovamente le fibre del fuso che agganciano i cinetocori dei cromosomi. Ai poli opposti della cellula si cominciano a formare i nuclei e avviene la citodieresi, con la conseguente scissione cellulare e i microtubuli del fuso scompaiono.
L'Ovogenesi: Peculiarità della Formazione dei Gameti Femminili
L'ovogenesi, detta anche oogenesi o gametogenesi femminile, è il processo mediante il quale si formano i gameti femminili, detti anche ovuli o ovociti. Talvolta si fa ancora riferimento ai gameti femminili chiamandoli uova, ma lo si usa in particolare solo per le specie animali. Per esempio, l'intero sviluppo prenatale degli uccelli ha luogo all'interno di un uovo. Nei mammiferi placentari, invece, dopo che l'uovo è stato fecondato e inizia a dividersi, nessuno lo chiama più uovo. Quindi, il termine più indicato per un gamete femminile maturo è ovulo (o ovuli, per la forma plurale). Questo processo avviene nelle ovaie, ovvero nella gonade femminile e si ripete con andamento ciclico (ciclo mestruale e in generale ciclo estrale per la maggior parte dei mammiferi placentati) per tutta la durata della fase di fertilità della femmina, che ha inizio con il menarca (la prima mestruazione) e termina con la menopausa.
Ovogenesi e Spermatogenesi: Un Confronto Rilevante
Il processo di formazione dei gameti si distingue in spermatogenesi per il maschio e ovogenesi per la femmina. Sebbene entrambi siano processi meiotici, l'ovogenesi differisce per molti aspetti dalla spermatogenesi. La principale differenza riguarda il numero di uova prodotte nel corso della vita di un individuo e la citodieresi.
In un maschio sano, i testicoli producono circa 100-200 milioni di spermatozoi al giorno. Della durata di circa 64 giorni, il processo di formazione e maturazione degli spermatozoi ha inizio nei tubuli seminiferi di ciascun testicolo e prosegue nell'epididimo. In assenza di eventi eiaculatori, gli spermatozoi hanno una vita limitata all'interno dei testicoli. Occorre precisare che la spermatogenesi è un processo continuativo, dotato di un proprio ritmo produttivo.
Nell'ovogenesi, invece, le primitive cellule germinali dell’embrione, i protogoni, sono simili nei due sessi. Per un certo periodo (moltiplicazione) esse si moltiplicano ripetutamente e danno origine a spermatogoni e ovogoni destinati a dare origine agli spermi e alle uova. Gli spermatogoni e ovogoni si dividono varie volte per mitosi, aumentando notevolmente di numero, e infine si differenziano in spermatociti e ovociti di primo ordine (detti anche primari).
La serie femminile presenta una peculiarità significativa: la maturazione degli ovogoni a ovociti primari avviene solo durante la vita intrauterina. Alla fine della gestazione, gli ovociti primari s’arrestano ai primi stadi della 1a divisione meiotica e rimangono quiescenti fino alla pubertà, quando si ha la riattivazione della meiosi. Le donne nascono con un ben preciso corredo di cellule uovo primitive.
Inoltre, lo scopo dell'ovogenesi è quello di produrre da ciascun oocita I una sola cellula uovo che contiene tutta la massa di citoplasma necessaria a sostenere lo sviluppo e il metabolismo dello zigote. Questa è una differenza cruciale rispetto alla spermatogenesi, dove da una singola cellula germinale si formano quattro spermatozoi funzionali. L'ovogenesi si caratterizza per la citodieresi ineguale che accompagna la prima e la seconda divisione meiotica, un meccanismo che assicura che quasi tutto il citoplasma sia conservato nell'ovulo.
Spermatogenesi
Le Fasi Chiave dell'Ovogenesi e l'Arresto Meiotico
Nell'uomo e in altri mammiferi, l'ovogenesi inizia durante la vita intrauterina. Le cellule germinali primordiali dell'epiblasto invadono per la prima volta il sacco vitellino nella terza settimana di sviluppo embrionale per proliferare per mitosi e nella settima settimana si spostano e invadono le ovaie differenziandosi in ovogoni. Il loro numero aumenta per mitosi, raggiungendo il massimo al settimo mese con un totale di 7 milioni di ovogoni, ma molti inizieranno a degenerare e ad atrofizzarsi. Gli esseri umani, infatti, nascono con un corredo ben preciso di cellule uovo primitive. In effetti, circa uno o due mesi prima della nascita di una bambina, all'incirca sette milioni dei suoi ovogoni muoiono, mentre i restanti ovogoni sopravvissuti entrano in meiosi I e diventano ovociti primari.
Questi ovociti primari mettono quindi in pausa il loro sviluppo fermandosi allo stadio di profase I della Meiosi, dopo aver replicato i loro genomi, ma prima di aver effettuato la prima divisione meiotica. Gli ovociti rimangono fermi in questa fase dello sviluppo per oltre 10 anni, fino a quando la ragazza ha il suo primo ciclo mestruale. Questi ovociti I prodotti resteranno "congelati" fino alla pubertà. Una donna nasce con 1 o 2 milioni di ovociti e al menarca si raggiunge la riattivazione dell'oogenesi con circa 400.000 ovociti, di cui un totale di 400-500 saranno ovulati fino alla menopausa.
A partire dalla pubertà, ogni mese, stimoli ormonali inducono la ripresa della meiosi di uno o pochi oociti. Gli ovociti primari riprendono la meiosi dove l'avevano interrotta e completano la prima divisione meiotica. Quando l'ovocita primario riesce finalmente a completare la sua prima divisione meiotica, divide i cromosomi in modo uniforme. Tuttavia, questa divisione è ineguale dal punto di vista citoplasmatico: l'ovocito di primo ordine si divide una prima volta in due cellule figlie, entrambe aploidi e diverse fra loro: una più grande, l'ovocito di secondo ordine, e un'altra più piccola, il primo globulo polare. La formazione di un corpo polare consente all'ovocita primario di ridurre il suo genoma della metà e conservare la maggior parte del suo citoplasma nell'ovocita secondario. Il corpo polare non è un ovocita funzionale, ma degenera e muore. Il primo globulo polare in genere non si divide, pur avendo un corredo cromosomico aploide formato da cromosomi fratelli, non è in grado di proseguire la meiosi e quindi di dividersi ulteriormente.
L'ovocita secondario ha ancora due copie di ciascun cromosoma, quindi, per diventare un ovulo completamente funzionante, deve essere sottoposto alla seconda divisione meiotica. Questa seconda divisione meiotica è cruciale ma avviene con un secondo blocco nella metafase della seconda divisione meiotica. Se non viene fecondato, l'ovulo viene espulso con le mestruazioni. Se la fecondazione avviene, l'ovocita secondario completa la meiosi II, producendo l'uovo maturo e un secondo globulo polare, anch'esso di piccole dimensioni.
Il Ciclo Ovarico e lo Sviluppo Follicolare
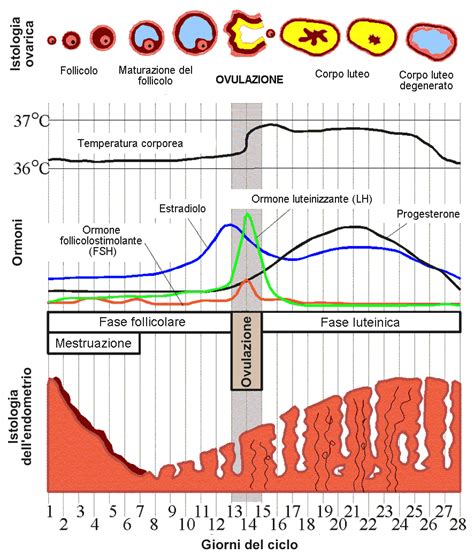
L'ovogenesi è strettamente integrata nel ciclo ovarico, che può suddividersi in tre momenti importanti: la fase follicolare, la fase ovulatoria (od ovulazione) e la fase luteale. Nell'uomo, lo sviluppo degli ovociti avviene nelle ovaie, dove si sviluppano i follicoli. L’oocita è contenuto nel follicolo ed è circondato da una serie di cellule follicolari che lo circondano e nutrono.
La fase follicolare è il momento in cui le ovaie, dopo un preciso stimolo ormonale, producono e portano a maturazione, all'interno di un follicolo, la futura cellula uovo. Al menarca, si raggiunge la riattivazione dell'oogenesi. A partire da questa età, ogni 4 settimane un follicolo viene portato a completa maturazione, insieme all'ovocita in esso contenuto. All'inizio del ciclo ovarico, il processo di maturazione interessa diversi follicoli, ma di norma soltanto uno raggiunge il completo sviluppo e viene espulso dall'ovaio per essere, eventualmente, fecondato. I rimanenti follicoli regrediscono rapidamente, secondo un processo degenerativo che interessa dapprima l'ovocita ed in seguito le cellule follicolari che lo circondano. Durante il processo di maturazione dell'ovocita, anche il follicolo subisce delle modificazioni, che lo portano a sostenere, dal punto di vista endocrino, la maturazione dell'ovulo in esso racchiuso.
L'azione contemporanea di FSH (Ormone Follicolo-Stimolante) e LH (Ormone Luteinizzante) stimola, inoltre, la secrezione di estrogeni da parte delle cellule follicolari. Gli estrogeni, a partire dal decimo giorno, inducono un picco di LH e un picco meno pronunciato di FSH. Questo picco di LH è il segnale per l'ovulazione.
La fase ovulatoria coincide generalmente con il mezzo del ciclo e cioè 14 giorni dopo l'inizio dell'ultima mestruazione. È il momento del rilascio della cellula uovo. A questo punto, l'ovulo avrà iniziato la seconda meiosi ma si fermerà nella metafase II. La maturazione in contemporanea di due o più ovociti è rara, ma comunque possibile. Se fecondati, questi ovuli possono dare luogo a due o più embrioni.
La fase luteale segue l'ovulazione. Dopo l'ovulazione, la cellula uovo viene prontamente catturata dalle fimbrie della tuba ed incanalata al suo interno. A questo punto il follicolo si trasforma in corpo luteo che secerne progesterone. Se avviene fecondazione, la produzione di progesterone continua. Se la cellula uovo non viene fecondata, entro una decina di giorni il corpo luteo cessa la produzione endocrina e regredisce, formando una piccolissima cicatrice sulla superficie ovarica (corpo albicante).
Il tempo che intercorre tra l'inizio dell'ovulazione e l'inizio della successiva mestruazione è pressoché costante (14 giorni perché gli eventi ormonali si controllano strettamente a cascata), il tempo necessario per portare l'ovulo a completo sviluppo e rilascio è molto variabile. L'ovocita ha una vita massima di 12-24 ore.
Morfologia dell'Ovocita
L'ovocita è una cellula molto complessa: in termini di dimensioni corrisponde a 100 micron metri, ossia un decimo di millimetro ed è in pratica la cellula di maggiori dimensioni dell’organismo. L'ovulo è una cellula di grandi dimensioni (100-150 micron di diametro), con il citoplasma ricco di materiali fosfolipidici di riserva (granuli di vitello o tuorlo). Questo citoplasma abbondante è essenziale per sostenere le prime fasi dello sviluppo embrionale.
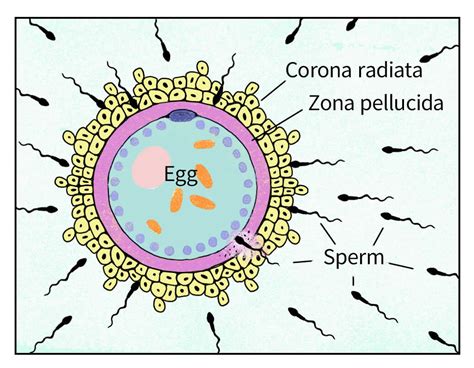
Dopo essere stato liberato nella cavità addominale, l'ovulo viene prontamente "risucchiato" dalla rispettiva tuba di Falloppio, sede deputata alla fecondazione. Affinché la fecondazione abbia luogo è necessario che uno spermatozoo penetri all'interno della cellula uovo. Si tratta di un evento delicato, dal momento che l'ovocita è protetto da alcune cellule (che costituiscono la cosiddetta corona raggiata) e membrane, come la zona pellucida, che si oppongono al loro ingresso. Per gli spermatozoi si tratta dunque di una vera e propria corsa ad ostacoli: solo il primo che riuscirà a raggiungere l'ovulo e a penetrarlo - grazie al rilascio di appositi enzimi - avrà l'onore di fecondarlo. In realtà - è bene puntualizzarlo - lo spermatozoo che penetra nella cellula uovo non agisce mai in solitario, ma si serve degli altri spermatozoi emessi con l'eiaculazione, per distruggere le barriere protettive esterne dell'ovocita.
Subito dopo la sua formazione, lo zigote si sposta dalle tube di Falloppio - sede canonica della fecondazione - all'utero. Dopo le prime divisioni, lo zigote assume il nome di blastocisti. Dopo circa 8 settimane dalla formazione dello zigote, la blastocisti diventa ciò che si chiama comunemente feto.
Errori Meiotici e le Loro Implicazioni nell'Ovocita
La meiosi, soprattutto nell'ovocita, è un processo sorprendentemente complesso e delicato, suscettibile a errori con conseguenze significative per la riproduzione e la salute dell'individuo. La meiosi sbaglia spesso. Quando una cellula uovo, o ovocita, matura all’interno dell’ovaio, va incontro ad una divisione cellulare chiamata meiosi nella quale le coppie di cromosomi si allineano e vengono “pescate”, cioè trasportate ai due poli opposti della cellula da filamenti proteici chiamati microtubuli.
La "Pesca" dei Cromosomi e la Suscettibilità agli Errori
Immaginate di osservare, ad una fiera di paese, un bambino con la fronte aggrottata per la concentrazione che sta cercando di pescare un pesce di legno da uno stagno di plastica, usando il magnete all’estremità della sua canna da pesca. Congelate la scena, riavvolgete tutte le tappe dello sviluppo del bambino, dopo il momento in cui lo sperma ha fecondato l’uovo, fino al punto in cui la cellula uovo si è formata, e troverete che anche nelle cellule avviene una sorta di pesca. Gli scienziati dell’EMBL, analizzando filmati dettagliati, hanno constatato che, prima che i microtubuli si attacchino ai cinetocori, spingono i cromosomi in una posizione favorevole, come un bambino che riposiziona un pesce con l’estremità della canna da pesca. Tuttavia, nonostante questo pre-posizionamento, l’operazione non funziona molto bene, rendendo la meiosi un processo intrinsecamente "imperfetto".
Questa "pesca" dei cromosomi comporta un maggior numero di errori nella meiosi rispetto alla mitosi. Durante la mitosi, i filamenti dei microtubuli iniziano a formarsi in due punti opposti della cellula e assumono man mano una forma a limone - il fuso - che successivamente trasporta (spinge) ciascun cromatidio del cromosoma verso uno dei due poli della cellula. Ma nella meiosi, come ha scoperto il gruppo di Jan pochi anni fa, i filamenti del fuso convergono partendo da almeno 80 punti diversi, e solo successivamente assumono una struttura a due poli. Per questo motivo è difficile sapere se i microtubuli che si attaccano ai cromosomi li spingeranno in direzioni opposte oppure no. Queste scoperte consentono agli scienziati di sapere dove guardare quando studiano l’infertilità femminile e le condizioni come la sindrome di Down, dovute in larga parte a cellule uovo con un numero di cromosomi anormale.
La Ricerca all'Avanguardia sugli Ovociti
Esaminando le cellule uovo di topo al microscopio, lo scienziato dell'EMBL Tomoya (Tomo) Kitajima è stato il primo a descrivere i movimenti di tutti i cinetocori di una cellula uovo durante la divisione cellulare - che dura circa 10 ore. Tomo ha usato un software - sviluppato nel laboratorio di Jan - che gli ha permesso di programmare un microscopio a scansione laser per individuare i cromosomi nel grande ambiente intracellulare della cellula uovo, e di filmarli durante la divisione. L’oocita è una cellula grande, ma i cromosomi si localizzano solo in una piccola parte di essa, e questo era ciò a cui eravamo interessati. Focalizzando il microscopio solo sulla parte della cellula contenente i cromosomi, Tomo ha potuto ottenere immagini ad alta risoluzione a brevi intervalli di un minuto e mezzo, che hanno fornito una dettagliata immagine del processo.
Dimostrando che questi errori in molti casi derivano dal mancato o errato aggancio dei microtubuli ai cromosomi, Tomo e Jan hanno fornito un riferimento per studi futuri. Infatti, Tomo sta ora studiando perché questo processo per tentativi ed errori è ancor più soggetto a sbagli nelle cellule uovo più vecchie. Se lui o altri riusciranno a capire dove il meccanismo di correzione fallisce nelle cellule più vecchie, questo potrebbe essere in futuro il punto di partenza per procedure mediche che aiutino i microtubuli a migliorare la tecnica di pesca, ovvero la corretta separazione dei cromosomi.
Il software che Tomo ha usato per trovare e filmare i cromosomi durante la divisione cellulare è stato il punto di partenza per ulteriori miglioramenti. Da allora, in collaborazione con un altro gruppo di ricerca che lavora all’EMBL, capitanato da Rainer Pepperkok, il gruppo di Jan ha sviluppato un programma più complesso, in grado di automatizzare ancor più il funzionamento. Una volta che Micropilot ha identificato la cellula o la struttura alla quale gli scienziati sono interessati, automaticamente dà istruzioni al microscopio, che possono essere semplicemente di registrare video ad alta risoluzione oppure utilizzare un laser per interferire con proteine rese fluorescenti e registrare i risultati. Questo software è molto utile negli studi di biologia dei sistemi perché produce un maggior numero di dati a maggior velocità. Il Laboratorio Europeo di Biologia Molecolare (EMBL) è uno degli enti di ricerca di eccellenza nel campo della ricerca di base delle scienze della vita. EMBL è internazionale, innovativo e interdisciplinare. La ricerca di Jan e Tomo mette in evidenza che la “pesca” dei cromosomi comporta un maggior numero di errori nella meiosi rispetto alla mitosi, un aspetto cruciale per comprendere le anomalie cromosomiche.