La vita, nella sua incredibile diversità, inizia da un singolo punto: lo zigote. Questo minuscolo ammasso di materiale genetico, visibile a malapena a occhio nudo, è il seme da cui germoglierà un intero organismo. Il viaggio da questa cellula primordiale a un essere vivente complesso e funzionante è un'epopea biologica affascinante, orchestrata dal fenomeno del differenziamento cellulare.
Il differenziamento cellulare è il processo attraverso il quale una cellula iniziale, come lo zigote, dà origine gradualmente a una moltitudine di tipi cellulari diversi, ognuno con la propria forma e funzione specifica, pur condividendo lo stesso identico patrimonio genetico. Questo prodigio biologico è alla base della pluricellularità e dell'evoluzione degli organismi complessi.
La Danza dei Geni: Programmazione Genetica al Cuore del Differenziamento
Alla radice del differenziamento si trova una sofisticata programmazione dell'attività genetica. In un primo tempo, questa programmazione determina la formazione di tipi cellulari distinti. Successivamente, assicura che queste cellule rimangano specializzate, mantenendo costantemente "represse" le sintesi genetiche che non sono specifiche del loro ruolo. Immaginate un'orchestra sinfonica: ogni musicista suona uno strumento diverso, ma tutti seguono la stessa partitura, eseguendo solo le note che appartengono alla loro sezione. Allo stesso modo, le cellule specializzate attivano solo i geni necessari alla loro funzione.
Questa capacità di modulare nel tempo e nello spazio l'attività dei vari gruppi di geni nucleari è stata probabilmente un motore fondamentale della grande fioritura evolutiva degli eucarioti. Molti di questi organismi, grazie a tale possibilità, sono diventati pluricellulari. Un batterio, il cui genoma è continuamente attivo, dà origine solo a cellule sempre eguali tra loro, che non traggono vantaggi dall'associazione perché tutte esercitano le stesse attività. Al contrario, l'organismo pluricellulare è composto da tipi cellulari diversi che, modulando in modo differente l'attività di singoli gruppi genici, si specializzano ciascuno per determinate funzioni, ricavando così vantaggi dalla collaborazione reciproca all'interno di un unico individuo.
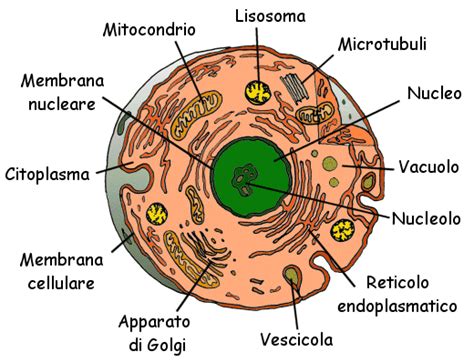
In un organismo adulto, le cellule dei vari tessuti differiscono tra loro per forma e funzioni. A livello molecolare, queste differenze si manifestano principalmente in proteine di carattere strutturale o enzimatico. La fibra muscolare, ad esempio, è enormemente più ricca di mioglobina, miosina, actina e altre proteine contrattili rispetto a qualsiasi altra cellula. Un globulo rosso, d'altro canto, non produce altro che emoglobina, la proteina essenziale per il trasporto dell'ossigeno.
Oltre il Nucleo: Il Ruolo Cruciale del Citoplasma e dei Segnali Esterni
È fondamentale comprendere che il nucleo non è l'unico responsabile di questa programmazione. Infatti, le cellule dei vari tessuti di un organismo possiedono tutte lo stesso DNA. La diversa regolazione dell'attività genetica che si attua nei vari tipi cellulari dipende da segnali chimici. Questi segnali possono giungere al nucleo dal citoplasma, oppure, sempre mediati dal citoplasma, da cellule circostanti o persino dall'ambiente esterno alla cellula.
Segnali chimici analoghi sono prodotti anche nel corso della normale vita cellulare, indipendentemente dal differenziamento. I complessi cicli vitali della cellula eucariotica sono programmati correttamente da molecole specifiche che vengono sintetizzate in determinati momenti e inducono il nucleo a iniziare una nuova fase di attività. Le interazioni nucleo-citoplasmatiche sono quindi alla base sia del differenziamento sia della normale attività di una cellula nel corso del suo ciclo vitale.
La Genesi dei Tipi Cellulari: Dalla Struttura dell'Uovo alla Specializzazione
La nascita dei vari tipi cellulari durante lo sviluppo embrionale dipende in larga misura dalla struttura molecolare della cellula-uovo. Fin dalla fecondazione, o anche prima in alcuni organismi, si osserva che il citoplasma ovulare non è omogeneo per quanto riguarda la distribuzione di determinati costituenti, in particolare le ribonucleoproteine (RNP). Si ritiene che queste RNP contengano RNA messaggeri temporaneamente inattivi, bloccati dagli informosomi.
Man mano che l'uovo si segmenta in numerosi blastomeri (cellule ancora indifferenziate), ciascun blastomero eredita una diversa porzione del citoplasma ovulare e, di conseguenza, differenti RNP. Successivamente, da questi blastomeri si originano cellule che si avviano in varie direzioni differenziatrici: alcune diventeranno muscolari, altre nervose, altre intestinali e così via.
Si ipotizza che questo processo sia dovuto a una diversa regolazione del loro DNA da parte di proteine citoplasmatiche particolari, costruite sui vari mRNA derivati dall'uovo e ora attivati.
Questi "controllori citoplasmatici" delle attività nucleari sono ancora in gran parte sconosciuti. Un esempio notevole è il fattore o+ (ova-deficient) scoperto nell'Axolotl, un anfibio che spesso non va incontro a metamorfosi. L'assenza di questa proteina provoca la produzione di uova che si arrestano nello sviluppo. Si ritiene che la proteina o+ sia necessaria per l'attivazione dei geni dei blastomeri, che altrimenti restano repressi. Infatti, uova segmentate di individui deficienti di questo fattore, destinate ad abortire, riprendono un normale sviluppo se viene loro iniettata la proteina o+, manifestando una ripresa della trascrizione nei nuclei dei blastomeri.
La segregazione di particolari RNP in determinate aree dell'uovo è provata anche da esperienze sui plasmi polari, zone ovulari ricche di granuli di RNA che normalmente danno origine alle gonadi dell'individuo. Se nell'uovo di una Drosofila vengono trapiantati granuli polari di un altro uovo, si origina un embrione con un duplice corredo di gonadi. Questi granuli contengono probabilmente mRNA di proteine che regolano il differenziamento delle cellule germinali.
La Plasticità Nucleare: Come il Citoplasma Riprogramma il DNA
Anche le esperienze di trapianto nucleare indicano che il citoplasma contiene molecole in grado di modificare le attività nucleari. È noto che il genoma di cellule differenziate trascrive pochi tipi di RNA, essendo in larga misura represso. Se il nucleo di una di queste cellule viene trapiantato in un uovo privato del proprio nucleo, nel nuovo ambiente il nucleo differenziato subisce una riprogrammazione: il DNA inizia ampi fenomeni di trascrizione e il nucleo è in grado di produrre il normale sviluppo dell'uovo, con la genesi di numerosi e diversi tipi cellulari. Per esempio, in una specie di anfibi si sono ottenuti individui completi iniettando nuclei di cellule intestinali in uova previamente enucleate.
I fenomeni regolativi dell'attività genetica sono dunque reversibili, in base a stimoli provenienti dal citoplasma. Un'ulteriore prova si ha da esperienze di fusione cellulare o ibridazione cellulare. La fusione di due cellule genera un ibrido con due nuclei in un unico citoplasma. Se le cellule appartengono a due specie diverse, l'ibrido è detto eterocarione. Sono stati ottenuti eterocarioni provocando la fusione di eritrociti di pollo (il cui nucleo è del tutto eterocromatico e quindi represso) con fibroblasti umani o di topo (il cui nucleo è invece molto attivo nella trascrizione). Nell'ibrido, il nucleo dell'eritrocita si ingrandisce rapidamente e la sua cromatina diviene attiva, trascrivendo RNA e fabbricando il nucleolo di cui era privo. Evidentemente, fattori presenti nel citoplasma del fibroblasto hanno attivato il DNA dell'eritrocita.
Ormoni e Puffs Cromosomici: Segnali Chimici e Attivazione Genica
Infine, è stato dimostrato che gli ormoni steroidei attivano alcuni tipi cellulari, quelli degli organi bersaglio, penetrando nel loro citoplasma e legandosi a recettori specifici di natura proteica. Questi recettori, attivati dallo steroide, passano nel nucleo ed entrano in rapporto con determinate regioni del genoma, sulle quali agiscono come derepressori, attivandone la trascrizione. Prove al riguardo si sono ottenute per gli steroidi sessuali (testosterone, estrogeni e progesterone) e della corteccia del surrene nei vertebrati, e per quelli della muta larvale negli Insetti (ecdisone).
In quest'ultimo caso, si è osservato che nei cromosomi politenici dei ditteri l'iniezione di ecdisone provoca la formazione di puffs (aree despiralizzate ad intensa attività trascrizionale) in determinate regioni di certi cromosomi, specifiche sia a livello di tessuto sia a livello dello stadio larvale osservato. Probabilmente, nei puffs più grandi, oltre a fenomeni di attivazione, avvengono anche fenomeni di amplificazione dell'attività genica.
L'amplificazione genica, ossia l'incremento numerico delle copie di un certo gene in attività, è stata recentemente indotta sperimentalmente anche in cellule di mammiferi in coltura. Trattando queste cellule con inibitori della sintesi delle purine (sintesi di vitale importanza), alcune di esse acquisivano gradualmente una resistenza all'agente inibitore, aumentando enormemente la quantità degli enzimi addetti alla sintesi delle purine, che quindi non venivano totalmente inibiti. A livello microscopico, si è osservato che queste cellule resistenti possedevano un cromosoma con un braccio enormemente più lungo della norma. Ci sono prove che lungo questo braccio, i geni che codificano l'enzima per la sintesi purinica siano presenti in alcune centinaia di copie, essendosi amplificati nel corso di varie generazioni sotto la pressione selettiva degli inibitori.
I principali eventi che caratterizzano le attività vitali della cellula sono dunque il risultato di interazioni, talora complesse, fra nucleo, citoplasma e ambiente.
Introduzione all'anatomia - cellule e differenziamento
La Struttura al Servizio della Funzione: Organuli Cellulari e Specializzazione
Il passo successivo, dopo aver compreso l'espressione genica, è rispondere alla domanda di come una cellula possa organizzare una struttura perfettamente in grado di espletare funzioni articolate. Si ritiene che ciò si realizzi grazie a un ordinato montaggio di macromolecole proteiche, lipidiche e zuccherine, secondo un preciso progetto strutturale che richiede l'intervento di attività enzimatiche in una sequenza temporale estremamente precisa.
Generalmente, l'acquisizione di una determinata funzione da parte di una popolazione cellulare si realizza quando in essa si è progressivamente accumulata una quantità considerevole di una o più proteine specifiche o proteine enzimatiche caratteristiche, in grado di espletare funzioni specializzate. Una caratteristica importante di alcune proteine del secondo gruppo è quella di non essere strettamente necessarie al metabolismo e, in particolare, alla sopravvivenza della cellula stessa. Pertanto, ai fini dell'espressione di attività cellulari caratteristiche, è necessario che ogni singola cellula sintetizzi non soltanto proteine in quantità sufficiente, ma anche rispettando una rigida sequenza temporale.
Durante il differenziamento, la cellula, in base alla qualità e alla disponibilità delle proteine prodotte, è in grado successivamente di assemblarle, acquisendo caratteristiche fenotipiche fortemente differenziate. Queste modificazioni interessano tutti i compartimenti cellulari, che possono tuttavia ulteriormente differenziarsi in rapporto alle funzioni specifiche che la cellula dovrà svolgere.
È importante sottolineare come la funzione finale svolta da ogni singola popolazione sia in gran parte dipendente dall'assortimento e dal numero dei macchinari molecolari presenti all'interno della cellula. In particolare, lo sviluppo dei vari compartimenti cellulari è responsabile di conferire talune caratteristiche attività funzionali.
Esempi di Specializzazione Strutturale: Dall'Intestino al Muscolo
È interessante osservare le differenze strutturali che i meccanismi differenziativi determinano a carico di alcuni tra i più importanti organuli cellulari.
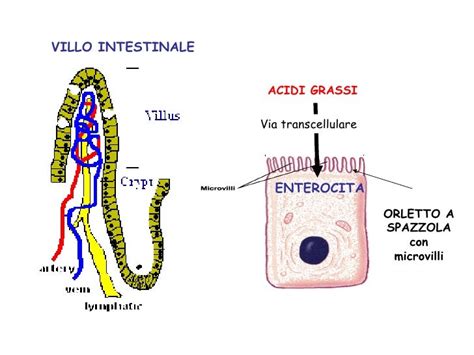
- Cellule Intestinali (Enterociti): Cellule che si sviluppano sulle pareti delle cavità di un organo, come l'intestino, possono specializzare la loro porzione apicale per lavorare in questa cavità, dove può essere presente materiale macro e micromolecolare di varia natura che deve essere assorbito, modificato o semplicemente analizzato. La porzione apicale delle cellule epiteliali dell'intestino, dette enterociti, riesce ad amplificare notevolmente la superficie cellulare attraverso una ripetuta pieghettatura che conferisce un aspetto a pettine.
Cellule della Cistifellea: Le popolazioni cellulari che rivestono il lume della cistifellea presentano microvilli con antenne, presumibilmente espressioni differenziate del "cell coat" che favoriscono il riassorbimento dell'acqua, importante ai fini della concentrazione della bile.
Cellule Tracheali Ciliate: Nella mucosa tracheale esistono cellule dotate di ciglia che, contraendosi, svolgono un lavoro meccanico di allontanamento nei confronti del materiale in transito.
Cellule Ciliate nei Condotti Riproduttivi: Molte volte la presenza di ciglia può addirittura promuovere o indurre il movimento di popolazioni cellulari, come accade nei condotti efferenti delle vie spermatiche maschili e degli elementi che rivestono la mucosa delle tube uterine. Grazie all'attività meccanica esercitata dalle ciglia, che battono in maniera orientata, ritmica e precisa, si creano dei moti propulsivi attivi in grado di determinare, rispettivamente, lo spostamento degli spermatozoi e dell'ovocellula.
Melanociti: Un ultimo esempio di collegamento fra struttura e funzione si trova nei melanociti, i quali, attraverso le propaggini citoplasmatiche denominate dendriti, possono mettersi in contatto con le cellule dell'epidermide per riversarvi il pigmento melanico. Durante il differenziamento di queste popolazioni cellulari, risulta evidente come debba essere fortemente incrementata la sintesi dei vari materiali molecolari che costituiscono queste strutture e come debba essere particolarmente controllato il loro successivo montaggio.
Attraverso la comparsa di caratteristiche fenotipiche fortemente specializzate, le varie popolazioni cellulari riescono a svolgere funzioni diverse e complesse.
Reticolo Endoplasmatico: Un Indicatore di Attività Sintetica
L'estensione del reticolo endoplasmatico rugoso (RER) è un criterio differenziativo in grado di fornire importanti informazioni sulle capacità di sintesi di una cellula.
Cellule Pancreatiche Esocrine: La cellula pancreatica presente nell'acino esocrino è un esempio di come sia possibile correlare una marcata attività secretoria con un imponente sviluppo del RER. L'elaborato di questa cellula andrà a costituire il succo pancreatico, una miscela di circa quaranta enzimi, che verrà immessa nella cavità duodenale per integrare l'attività intestinale nella demolizione delle molecole presenti nel chimo.
Epatociti e Reticolo Endoplasmatico Liscio: Il reticolo endoplasmatico liscio (REL) dell'epatocita può variamente ampliarsi durante tutto il corso della vita in seguito all'azione di sostanze eterogenee, tra cui i farmaci. È stato calcolato che, in seguito a somministrazione di barbiturici, il REL può ipertrofizzarsi, raggiungendo valori pari a circa 45.000 mm² (fenomeno noto come farmacoinduzione). Il REL è senza dubbio la struttura più enigmatica dello sviluppo in quanto non è possibile dedurre la funzione della cellula basandosi solamente sull'analisi della sua organizzazione strutturale. Durante la citodifferenziazione, il REL, scarso e poco delineato nelle cellule pluripotenti, tende a svilupparsi costituendo complicati sistemi.
L'Inizio della Vita: Lo Sviluppo Embrionale Umano
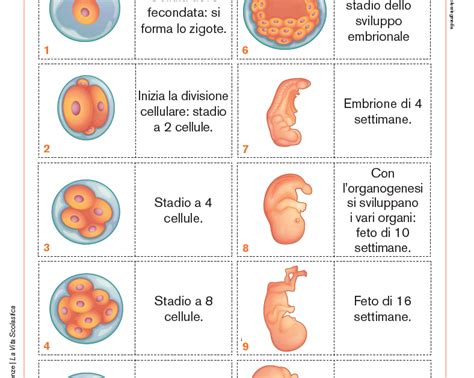
Ma come si arriva a questo stadio di complessità partendo da una singola cellula? L'embriologia, la scienza che studia i processi di nascita, crescita e sviluppo degli organismi viventi, ci offre la risposta. Nel caso della specie umana, tutto parte dalla fecondazione, il processo in cui la cellula uovo femminile riceve al suo interno uno spermatozoo maschile. La fusione di questi due gameti genera lo zigote, l'inizio della nostra vita. Dopo circa 40 settimane, da questa singola cellula si svilupperà un bambino con tutti gli organi formati e pronti per affrontare il mondo extrauterino.
Vediamo brevemente le tappe fondamentali:
- Fecondazione: La cellula uovo e lo spermatozoo si fondono, formando lo zigote.
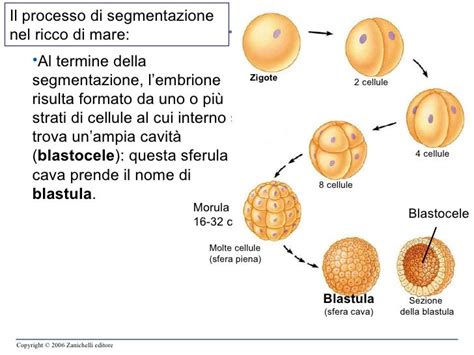
- Segmentazione: Dopo circa 24 ore, lo zigote inizia a dividersi ripetutamente tramite mitosi, dando origine a cellule sempre più piccole chiamate blastomeri.
- Morula: Dopo tre giorni, si forma la morula, costituita da 8-16 blastomeri.
- Blastula e Gastrulazione: Dalla morula si passa alla blastula, costituita da numerosi blastomeri. È in questo momento che avviene il processo di differenziamento cellulare. Le cellule si riuniscono a seconda del loro scopo finale, formando tre foglietti embrionali sovrapposti: l'ectoderma (esterno), il mesoderma (intermedio) e l'endoderma (interno). Questo processo è chiamato gastrulazione ed è fondamentale perché ogni cellula ha un compito specifico e deve collocarsi nel foglietto embrionale corretto affinché ogni organo si formi in maniera appropriata.
- Organogenesi: Grazie alla presenza dei tre foglietti embrionali, si iniziano a formare gli organi e i tessuti, che giungeranno a completamento, in genere, poco prima della nascita.
Possiamo distinguere due periodi principali di sviluppo intrauterino:
- Periodo Embrionale (3-8 settimane): In questa fase, il differenziamento cellulare dà origine alle varie parti del corpo e ai tessuti (si formano testa, cervello, midollo spinale, occhi, orecchie, naso, sistema nervoso, reni, polmoni, ecc.). L'embrione sviluppa sembianze umane riconoscibili e viene chiamato feto. Alla fine dell'ottava settimana, l'embrione misura circa 2,5 cm.
- Periodo Fetale (9 settimane - nascita): In questo lungo periodo, si conclude lo sviluppo fisico. La testa del feto, inizialmente grande quanto il resto del corpo, si ridimensiona. A partire dal terzo mese, gli organi si sono formati e iniziano a funzionare (ad eccezione dei polmoni, che si svilupperanno completamente ma non saranno funzionali fino alla nascita, essendo pieni di liquido amniotico). Dal quarto mese, la madre percepisce i movimenti del bambino, e questo è il periodo di crescita più veloce. Dal quinto mese si sviluppano pelle, capelli e unghie. Dal sesto mese, il feto può aprire e chiudere gli occhi e distinguere luce da buio. Nell'ultimo trimestre, il feto si posiziona generalmente a testa in giù ("posizione cefalica") per prepararsi al parto naturale.
Determinazione del Sesso: Un Complesso Equilibrio Genetico e Ormonale
Un aspetto cruciale dello sviluppo è la determinazione del sesso, un processo che porta all'acquisizione differenziale di caratteri sessuali maschili e femminili, fondamentale per la riproduzione e il mantenimento della specie. Questo processo avviene attraverso meccanismi complessi che coinvolgono geni, cromosomi e ormoni.
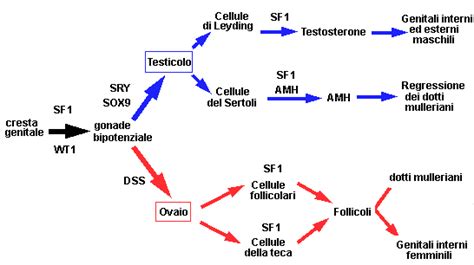
Nei mammiferi, il sesso è determinato principalmente dal sistema XX/XY. Le femmine possiedono due cromosomi X (XX), mentre i maschi hanno un cromosoma X e uno Y (XY). Il cromosoma Y porta il gene SRY (Sex-determining Region Y), considerato il "gene interruttore" che avvia lo sviluppo del testicolo.
Le gonadi (testicoli e ovaie) si formano nell'ambito del sistema uro-genitale in via di sviluppo, che origina dal mesoderma intermedio. Il primordio gonadico, inizialmente identico in entrambi i sessi, è costituito da uno strato superficiale (cortex) e una regione interna (medulla).
- Sviluppo Testicolare (XY): In presenza del gene SRY, il primordio gonadico si differenzia rapidamente in testicolo. SRY attiva a cascata una serie di geni, tra cui SOX9, che gioca un ruolo chiave nello sviluppo testicolare. SOX9, a sua volta, promuove l'espressione di altri geni, come FGF9, che creano un circuito di feedback positivo mantenendo l'espressione di SOX9 e inibendo al contempo i geni coinvolti nello sviluppo ovarico (come WNT4). Le cellule del Sertoli, che si differenziano all'interno dei futuri tubuli seminiferi, producono l'ormone anti-Mülleriano (AMH), che causa la degenerazione dei dotti di Müller (destinati a formare le vie genitali femminili), e il testosterone, che promuove lo sviluppo dei dotti di Wolff (destinati a formare le vie genitali maschili).
- Sviluppo Ovarico (XX): In assenza di SRY, e in presenza di due cromosomi X, il primordio gonadico si sviluppa in ovaio. Questo processo è mediato dall'azione di più geni, tra cui WNT4 e FOXL2, che promuovono lo sviluppo ovarico e inibiscono i pathway maschili. L'espressione di SOX9 viene repressa, mentre i dotti di Müller persistono e i dotti di Wolff degenerano.
La determinazione sessuale è un processo finemente regolato, sensibile alla dose genica e al tempo di espressione. Mutazioni o alterazioni in questi geni possono portare a disordini dello sviluppo sessuale, come il pseudoermafroditismo.
La Scoperta di Ell3: Un Fattore Chiave nell'Attivazione Genica Rapida
Una scoperta recente ha gettato nuova luce sui meccanismi che permettono alle cellule staminali di passare rapidamente da uno stato di pluripotenza a una specifica linea cellulare, un processo di differenziazione che richiede la rapida e contemporanea attivazione di centinaia di geni. Si tratta di una proteina chiamata Ell3, prodotta da un gene (Eleven-nineteen lysine-rich leukemia gene) la cui funzione era stata finora sottovalutata.
Ell3 agisce come un fattore di allungamento, incrementando la velocità con cui i geni vengono espressi. Essa si posiziona in corrispondenza del sito di attacco per le proteine attivatrici (enhancer) dei geni, reclutando la RNA polimerasi II. Questa enzima, che provvede alla traduzione delle istruzioni codificate nel DNA, rimane così in uno stato di "pausa" ma con i "motori accesi", pronta a svolgere la propria funzione non appena arriva il segnale di iniziare la differenziazione.
La proteina Ell3 è stata identificata come fattore di allungamento nel 2000, ma si sapeva solo che era altamente espressa nei testicoli, suggerendo un suo possibile intervento nello sviluppo embrionale. Ricerche recenti su cellule staminali embrionali di topo hanno rivelato che Ell3 si posiziona in corrispondenza di oltre 5000 enhancer, molti dei quali regolano geni che controllano la maturazione di cellule staminali di midollo spinale, renali ed ematiche. Questa scoperta è rilevante non solo per la biologia dello sviluppo, ma anche per l'oncologia, dato che molti di questi stessi geni risultano anormalmente attivi in diverse forme tumorali.
Induzione Embrionale e Campi Morfogenetici: Comprendere l'Organizzazione dello Sviluppo
Lo studio della regolazione dei processi di sviluppo ha rivelato complicate dinamiche interattive tra le parti dell'embrione. Il sorprendente fenomeno dell'induzione embrionale, descritto da Hans Spemann e Hilde Mangold nel 1924, dimostrava cambiamenti di potenzialità e competenze dei tessuti durante le diverse fasi dello sviluppo. Spemann e Mangold verificarono che una regione specifica dell'embrione allo stadio di gastrula, il labbro dorsale del blastoporo, se trapiantata su un altro embrione, aveva la capacità di riorganizzare le cellule della regione ospite inducendo la formazione di un nuovo embrione.
La fenomenologia dell'induzione dimostrava che diverse regioni dell'embrione hanno la potenzialità di indurre la formazione di strutture nelle regioni vicine, le quali, tuttavia, devono possedere la competenza necessaria per poter essere indotte a cambiare destino.
L'individuazione dei principi generali dello sviluppo è stato l'obiettivo centrale dell'embriologia sperimentale. Tuttavia, fino agli anni Cinquanta, questa disciplina fu dominata dall'idea che la vita fosse governata da "leggi della forma". Gli embriologi osservavano il comportamento "olistico" dell'embrione o di alcune sue regioni, trovando inverosimile che una teoria "discontinuista" come quella evoluzionistica, basata sul controllo genetico di caratteri fenotipici definiti, potesse spiegare la fenomenologia embrionale con i suoi aspetti di continuità e plasticità.
Si immaginò l'esistenza di campi morfogenetici, regioni transitorie dell'embrione in grado di autodifferenziarsi come un tutto, e in cui ogni parte poteva essere sostituita da altre, con un frammento di adeguate dimensioni capace di ricostituire il tutto. Questo concetto, analogo ai campi elettrici o magnetici descritti dalla fisica, fu utilizzato anche da Spemann, sebbene con scetticismo.
Con la scoperta del codice genetico e delle regole di traduzione del linguaggio basato sui nucleotidi in quello delle proteine, i biologi pensarono di possedere finalmente gli strumenti concettuali per spiegare anche la costruzione del fenotipo dall'informazione contenuta nell'uovo fecondato. I limiti di questo approccio emersero chiaramente dall'esito di un importante programma di ricerca avviato da Sidney Brenner, che si pose l'obiettivo di scoprire il "programma genetico" che coordina la regolazione, nello spazio e nel tempo, dei geni per costruire l'organismo completo. Utilizzando il verme Caenorhabditis elegans, Brenner riuscì a stabilire la genealogia di ciascuna delle 959 cellule che si sviluppano nell'animale adulto.
Il differenziamento cellulare è, dunque, un processo straordinario che trasforma una singola cellula in un organismo complesso, un viaggio affascinante guidato da una precisa programmazione genetica e da un'intricata rete di segnali molecolari. È la danza della vita, che inizia dal singolo e si espande nella meraviglia della diversità.
tags: #differenziamento #cellulare #da #embrione