La complessità intrinseca delle cellule riproduttive femminili, gli ovociti, rappresenta un campo di studio fondamentale per comprendere i meccanismi alla base della vita e dello sviluppo embrionale. Al centro di questa complessità vi sono strutture citoplasmatiche e processi metabolici dedicati alla preparazione dell'ovocito per la fecondazione e il successivo sviluppo. Tra queste, il "corpo vitellino" assume un ruolo di primaria importanza, fornendo le riserve nutritive essenziali. La comprensione di tali strutture e dei processi ad esse correlati ha beneficiato enormemente del lavoro di pionieri dell'istologia e dell'embriologia, tra cui figura Édouard-Gérard Balbiani, il cui contributo ha illuminato aspetti cruciali della biologia cellulare riproduttiva.
1. Il Vitello e il Concetto di Corpo Vitellino
Il vitello rappresenta la principale riserva nutritiva che sarà utilizzata dall'embrione durante le prime fasi del suo sviluppo, prima che l'organismo diventi autonomo o che si stabilisca una connessione efficiente con l'organismo materno. Questa riserva è cruciale per la sopravvivenza e la crescita dell'embrione, fornendo tutti i nutrienti necessari. Il vitello non è un'entità omogenea, ma una miscela complessa di diverse sostanze. È composto principalmente da lipidi, glicoproteine e albumina. Questi componenti sono organizzati in strutture specifiche all'interno dell'ovocito. Tra queste strutture, i componenti vanno ad applicarsi all'interno della membrana cellulare, formando depositi organizzati. È interessante notare come il vitello possa presentare una consistenza non è ben nota, o almeno la sua precisa organizzazione subcellulare. Si possono distinguere, ad esempio, granuli più piccoli dotati di un diametro di 2 mm, che contribuiscono alla sua complessa architettura. L'intera riserva nutritiva, spesso identificata come "corpo vitellino" o semplicemente "vitello," svolge un ruolo insostituibile nel sostenere l'inizio della vita. Nell'uovo in toto, si osserva una tipica organizzazione macroscopica in quanto non è fisicamente omogeneo, mostrando strati distinti di tuorlo di diverso colore e composizione. La sua organizzazione non è ben nota in tutti i dettagli, ma è evidente il suo ruolo di deposito energetico e strutturale.
2. La Vitellogenesi: Processi di Formazione del Vitello nell'Ovocito
Il processo di accumulo del vitello all'interno dell'ovocito è noto come vitellogenesi. Questo processo è una fase critica della maturazione ovocitaria, in cui l'ovocito si prepara per sostenere lo sviluppo embrionale. Durante la vitellogenesi, le strutture legate alla sintesi proteica si fanno più evidenti all'interno dell'ovocito. Questo indica un'intensa attività metabolica e di sintesi necessaria per produrre le grandi quantità di proteine e lipidi che compongono il vitello. Nel corso di questo processo, si formano numerosi vacuoli forniti di membrana, che servono a contenere e organizzare il materiale vitellino.
La formazione del vitello non avviene esclusivamente all'interno dell'ovocito stesso. Gran parte dei suoi costituenti, in particolare i precursori proteici, vengono sintetizzati altrove, come nel fegato. Questi precursori penetrano nell'oocita con modalità non ancora precisate, attraverso meccanismi di trasporto attivo e endocitosi. Una volta all'interno, subiscono una trasformazione delle vescicole citoplasmatiche circondate da membrana, che poi si fondono o si modificano per formare le riserve di vitello. Questa sintesi è continua per un certo periodo, assicurando un accumulo adeguato. L'organizzazione macroscopica del vitello nell'uovo non è fisicamente omogeneo, ma presenta bande di colore diverso (giallo scuro e giallo chiaro) che indicano fasi alterne di deposizione durante il giorno e la notte. I solidi presenti nel tuorlo sono rappresentati principalmente da lipidi, glicoproteine e albumina. Questi costituiscono la parte essenziale del vitello, fornendo l'energia e i blocchi costruttivi necessari per l'embrione. Il compito dei testi di fisiologia è quello di descrivere in dettaglio i meccanismi e la regolazione di questo processo vitale.
Gametogenesi, Fecondazione e Sviluppo dell'Uovo: Lezione Completa per il Test di Medicina 2025
3. L'Ovocito: Struttura, Sviluppo e Maturazione
Le cellule riproduttive femminili, gli ovociti, si formano prima della nascita, durante lo sviluppo degli organi genitali, a differenza di quelle maschili (spermatozoi). Questo significa che una donna nasce con una riserva ovarica predeterminata, composta da 1-2 milioni di follicoli. Tuttavia, alla pubertà ne rimangono circa 500.000. Nel corso della vita, questa "riserva ovarica" si riduce progressivamente ogni mese fino ad esaurirsi del tutto, segnando la comparsa della menopausa.
La maturazione ovocitaria è un processo altamente regolato. Un primo livello di selezione si basa sulla maturità nucleare ovocitaria. Al momento del prelievo ovocitario la maggior parte degli ovociti si trova invece ad uno stadio di maturazione ottimale per la fecondazione, ovvero Metafase II. Tuttavia, una percentuale compresa tra il 30 e il 70% degli ovociti Metafase I matura in-vitro e raggiunge in alcune ore lo stadio di Metafase II. Questi ovociti maturati in-vitro hanno ridotte percentuali di fecondazione ma simile qualità embrionaria rispetto agli ovociti Metafase II.
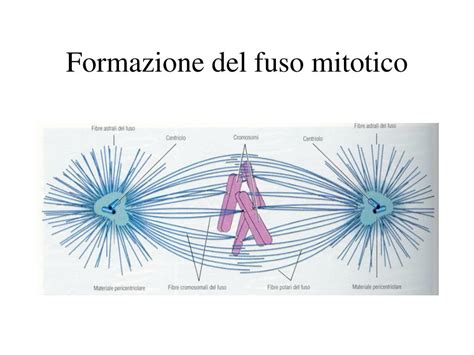
Oltre alla valutazione dello stato meiotico, è importante andare a valutare l’aspetto morfologico del citoplasma ovocitario, in quanto spesso circa la metà degli ovociti prelevati presentano almeno un’anomalia morfologica. L’integrità del fuso meiotico negli ovociti in Metafase II è di fondamentale importanza per una normale fecondazione ed un corretto sviluppo embrionario. Il fuso mitotico o meiotico è l’apparato cellulare che permette la divisione dei cromosomi nel corso della mitosi e della meiosi. La divisione cellulare si verifica a causa della bipolarità del fuso, costituito da due gruppi di polarità opposta, disposti intorno ai cromosomi. I limiti di questo criterio sono insiti nella struttura del fuso meiotico che risulta essere altamente sensibile a modificazioni chimico-fisiche che si possono verificare durante la manipolazione ovocitaria.
Quando l'ovocita primario completa la sua prima divisione meiotica, i cromosomi si suddividono in modo uniforme ma questo non avviene con il citoplasma. Il citoplasma si concentra per la maggior parte in una delle due cellule figlie (chiamata ovocita secondario) e rimane in piccole quantità nell’altra che viene chiamata corpo polare. Il corpo polare non è un ovocita funzionale, ma degenera e muore.
4. La Selezione Ovocitaria e i Criteri di Valutazione
Il sistema riproduttivo femminile dipende dal ciclico reclutamento follicolare, dalla selezione di un unico follicolo dominante, dall’ovulazione e dalla formazione del corpo luteo. Per ottimizzare le procedure di riproduzione assistita, la selezione degli ovociti è cruciale. L’entità della vascolarizzazione follicolare può rappresentare un importante parametro clinico di selezione follicolo-ovocitaria. La possibilità di identificare i follicoli maggiormente vascolarizzati da cui prelevare gli ovociti permetterebbe di ridurre il numero di embrioni da trasferire, riducendo al massimo il rischio di gravidanze multiple senza diminuire i tassi di gravidanza.
La tecnica di selezione ovocitaria basata sull’analisi cromosomica del primo corpo polare si basa sul principio secondo il quale tale corpo costituisce un'immagine speculare dell'ovocita. Una volta prelevato, il primo corpo polare viene trattato specificamente per poterne analizzare il materiale genetico. L'analisi genetica del primo corpo polare, se confrontata con quella eseguita sull'embrione, ha il vantaggio di consentire la selezione degli ovociti prima della fecondazione e inoltre non comporta la selezione di embrioni che presentino anomalie cromosomiche.
Secondo la Legge 40, la selezione ovocitaria deve essere quella dei 3 migliori ovociti da inseminare in base ai soli criteri clinici e morfologici, la cui correlazione con gli esiti clinici è ancora da dimostrare. Da un punto di vista clinico, sono incoraggianti i risultati riportati dalla ricerca sulla correlazione tra vascolarizzazione del follicolo e potenziale di sviluppo dell'ovocita corrispondente. Follicoli maggiormente vascolarizzati conterrebbero ovociti con maggiore potenziale di fecondazione e di sviluppo embrionario. Recentemente, la possibilità della valutazione del fuso meiotico ha offerto inoltre un altro criterio di selezione ovocitaria, come evidenziato da studi quali "Criteri morfologici di selezione ovocitaria" di Ubaldi FM, Rienzi L, Minasi MG, Greco E per la Società Italiana della Riproduzione.
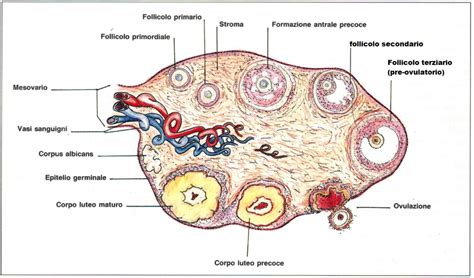
5. Le Ovaie: Ambiente di Sviluppo e Funzioni
Le ovaie costituiscono le gonadi femminili e svolgono una doppia funzione essenziale per la riproduzione. Queste funzioni sono: la funzione riproduttiva, che include la maturazione dei gameti, e la funzione endocrina, che si traduce nella produzione di estrogeni, progesterone e androgeni. Questi ormoni sono cruciali per la regolazione del ciclo riproduttivo e per il mantenimento della gravidanza.
L'ovaio è rivestito da un epitelio di rivestimento, di tipo cubico semplice, e da una falsa albuginea, che è un tessuto connettivo denso. La regione periferica dell'ovaio è costituita da stroma e organuli ovarici. Lo stroma è un tessuto connettivo particolarmente ricco di cellule e, al contempo, povero di fibre collagene. Questo stroma è, in particolare, un tessuto connettivo ricco di fibrociti capaci di trasformarsi in cellule secernenti ormoni steroidei, sottolineando la sua importanza nella funzione endocrina dell'organo. La riserva ovarica, come già menzionato, è un patrimonio limitato che si riduce progressivamente nel tempo.
Il sistema riproduttivo femminile è un meccanismo finemente orchestrato che dipende da una serie di eventi ciclici. Questi includono il reclutamento follicolare, la selezione di un unico follicolo dominante, il processo di ovulazione e, infine, la formazione del corpo luteo. Ogni donna nasce con 1-2 milioni di follicoli e alla pubertà ne rimangono 500.000. L'ovaio è un organo vitale, ed è la gonade femminile.
6. Édouard-Gérard Balbiani: Un Pioniere dell'Istologia e dell'Embriologia
Nell'ambito dello studio delle strutture microscopiche degli organismi e del loro sviluppo, la figura di Édouard-Gérard Balbiani si distingue come un pioniere. Istologo francese, Balbiani nacque a S. Domingo nel 1825 e morì a Meudon nel 1899. La sua carriera scientifica fu notevole e ricca di contributi significativi. Si addottorò in medicina a Parigi nel 1854, e poco dopo iniziò a pubblicare importanti memorie sui fenomeni sessuali negl'Infusorî. La qualità e l'originalità di queste ricerche furono tali che Claude Bernard, uno dei più grandi fisiologi del tempo, lo nominò chef des travaux d'histologie nel laboratorio di fisiologia generale del Museum. Balbiani mantenne questa prestigiosa carica fino al 1873, anno in cui fu chiamato a sostituire Coste nella cattedra di embriologia comparata al Collège de France, un'istituzione di altissimo livello accademico.
La sua influenza si estese anche alla fondazione di importanti pubblicazioni scientifiche. Insieme a Ranvier e Henneguy, Balbiani fondò gli "Archives d'Anatomie microscopique", una rivista che divenne un punto di riferimento per gli studi in istologia. Nel corso della sua vita, pubblicò numerose e importanti ricerche d'embriologia, d'istologia e d'anatomia. Il suo lavoro pionieristico nell'osservazione microscopica e nella descrizione dettagliata delle strutture cellulari e dei processi di sviluppo fu fondamentale. Sebbene il "corpo vitellino di Balbiani" non sia un termine uniformemente accettato come una struttura specifica da lui nominata, il suo estensivo lavoro in istologia ed embriologia, unito alla sua meticolosa analisi delle cellule germinali e dei primi stadi di sviluppo, ha senza dubbio contribuito in modo sostanziale alla comprensione delle inclusioni citoplasmatiche degli ovociti, inclusi i depositi vitellini e le loro dinamiche. Il suo approccio metodologico e la sua dedizione all'anatomia microscopica hanno posto le basi per successive scoperte e per una più profonda conoscenza di queste complesse strutture vitali per lo sviluppo embrionale.
7. Regolazione Endocrina e Ambientale della Riproduzione
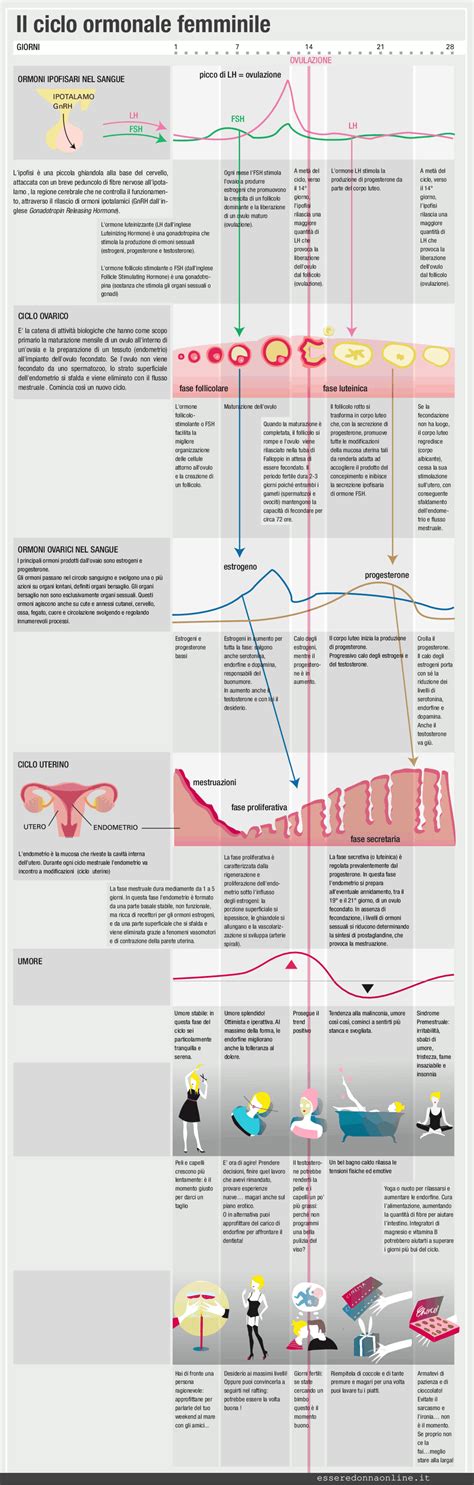
Il ciclo riproduttivo e la deposizione delle uova sono processi finemente regolati da un complicato insieme di fattori endogeni ed esogeni. Tra i fattori endogeni, gli ormoni sessuali giocano un ruolo preponderante. L'ipotalamo e l'ipofisi costituiscono un asse di controllo centrale, dove l'ipotalamo rilascia ormoni che stimolano l'ipofisi a produrre gonadotropine come l'FSH (follicolostimolante) e l'LH (luteinizzante).
L'FSH stimola la crescita e lo sviluppo dei follicoli, mentre l'LH è essenziale per l'ovulazione. Esistono due centri funzionali distinti nella regolazione ormonale. Gli estrogeni sono fondamentali perché si abbia un adeguato rilascio di LH, e l'LH stesso è l'ormone che consente l'ovulazione. La maturazione finale del follicolo nell'animale che si approssima alla prima ovulazione è accompagnata da un incremento del testosterone circolante, che contribuisce al processo complessivo. La deposizione è arrestata dalla prolattina, mentre l'LH ne è il principale stimolo. Il peduncolo dell'ovaio diventa ciclica con la maturità sessuale.
I fattori esogeni, in particolare il fotoperiodo, cioè le ore di luce giornaliere, esercitano un'influenza significativa. L'incremento del fotoperiodo funziona da stimolante per lo sviluppo delle gonadi, mentre il suo accorciamento funziona da inibente. Questo è particolarmente evidente in animali selezionati da soggetti viventi ai tropici. Un incremento del fotoperiodo può anticipare il raggiungimento della maturità sessuale, mentre la sua riduzione è in grado di ritardarla. Anche i regimi fotoperiodici interferiscono con la carriera riproduttiva nel suo complesso, influenzando sia l'inizio che la durata della vita produttiva.
La deposizione delle uova segue un ritmo quotidiano e ogni ciclo possiede una pausa. In razze allevate per la produzione di uova, la deposizione si verifica solitamente nel pomeriggio. Se una gallina depone un uovo nel pomeriggio, il prossimo uovo potrebbe essere deposto il giorno successivo, ma più tardi, oppure potrebbe saltare un giorno e deporre la mattina di dopodomani. Un anticipo di mezza giornata sull'ora della deposizione è possibile. Se la deposizione viene ritardata di oltre 24 ore, l'ovulazione successiva si verificherà durante la notte. L'iniezione di prostaglandina può anticipare la deposizione. Le contrazioni uterine, indotte anche dall'ossitocina, cui consegue la deposizione, sono stimolate da ormoni come l'arginina-vasotocina e la mesotocina, che stimolano i muscoli lisci e sono presenti nel sangue appena prima della deposizione. Questi ormoni agiscono sulla vagina dove inducono il rilasciamento della muscolatura, facilitando il passaggio dell'uovo verso la cloaca.
8. L'Ovidutto e la Formazione dell'Uovo Finale
L'ovidutto è una parte fondamentale del sistema riproduttivo femminile, responsabile della formazione delle varie componenti dell'uovo intorno all'ovocita dopo l'ovulazione. Nella gallina, questo aspetto riveste scarsa importanza dal punto di vista della presenza di un ovaio destro vestigiale; solo l'ovaio sinistro è funzionale. L'ovidutto è un lungo tubo diviso in diverse sezioni: infundibolo, magnum, istmo, utero e vagina.
L'infundibolo rappresenta l'estremità craniale dell'ovidutto ed è la sezione responsabile della raccolta degli oociti al momento dell'ovulazione. Le sue pareti sono spesse e descrive diverse anse. Il magnum è la parte più lunga dell'ovidutto (VIII. di circa 33 cm) ed è il sito dove viene secreto la maggior parte dell'albume. Qui, l'albume avvolge il tuorlo e lo protegge. L'istmo, la sezione successiva, misura circa 10 cm ed è dove si formano le membrane testacee, due strati che conferiscono resistenza all'uovo.
L'utero (o ghiandola del guscio), che misura circa 10 cm e ha un volume di 16-17 cm³, è il sito dove si forma il guscio calcareo esterno dell'uovo. Questa sezione presenta una mucosa caratterizzata da pliche secondarie e dalla presenza di ghiandole tubulari diffuse in tutto l'ovidutto. Gli epiteli secernenti e ciliati sono responsabili della produzione di muco e albume, che favorirebbero la progressione degli spermatozoi. Il guscio, per la sua notevole resistenza, specialmente verso la regione appuntita dell'uovo, è fondamentale per la protezione dell'embrione. Durante il passaggio attraverso l'ovidutto, l'albume si arricchisce di zuccheri e minerali.
Infine, la vagina, che misura circa 5 cm, è il tratto terminale dell'ovidutto che si apre nella cloaca. Il passaggio dell'uovo attraverso la vagina avviene in circa un minuto. La fisiologia della deposizione dell'uovo e i meccanismi ormonali precisi che la regolano non sono ben noti, sebbene sia chiaro il ruolo degli ormoni sessuali. Gli studi indicano che l'ipotalamo ne anticipa il rilascio, mentre l'esposizione allo stress lo ritarda.
Le lamine mucose che si incontrano nell'ovidutto, in particolare a livello dell'albume, sono essenziali per la formazione di una struttura completa. Queste lamelle, alte alcuni millimetri, sono rivestite di mucine che conferirebbero l'aspetto di gel denso all'albume. Le mucine si legano mediante mucine alle membrane testacee, rafforzando la struttura. La membrana esterna è più complessa della interna. All'interno dell'uovo, sono presenti le calaze, che sono strutture cordoniformi avvolte su se stesse, intimamente ritorte, formate da albume denso e che servono a mantenere il tuorlo centrato all'interno dell'albume.
9. Anomalie Morfologiche dell'Uovo
Il processo di formazione dell'uovo, sebbene altamente efficiente, è suscettibile a diverse anomalie morfologiche. La teratologia dell'uovo, che in greco significa "mostruosità", studia queste variazioni. Queste anomalie possono presentarsi in diverse forme e dimensioni, alcune delle quali sono piuttosto rare, mentre altre si osservano con una certa frequenza in diverse specie, dalla gallina della giungla all'anatra, oca, tacchina e struzzo.
Una delle anomalie più comuni è l'uovo senza tuorlo, o "avitellino". Questi piccoli uova, spesso descritte come "parvum, totum rotundum ad modum sperae" (piccolo, tutto rotondo come una sfera) da Aldrovandi nel XVI secolo, o come "nisi albugo, et fuit ovum" (solo albume, era un uovo) non contengono un tuorlo e sono costituite principalmente da albume. Di solito, l'uovo avitellino è più piccolo del normale per grandezza.
Un'altra anomalia è l'uovo con doppio tuorlo, che si verifica quando due ovociti vengono ovulati quasi contemporaneamente e vengono inglobati nello stesso albume. Questo porta a un uovo con doppio tuorlo. Se questo uovo rimane più a lungo in utero, può rivestirsi di ulteriore guscio, aumentando la sua dimensione e alterando la sua forma.
Le uova doppie, o uova con due gusci, sono un'anomalia interessante. In questi casi, un uovo completo viene deposto all'interno di un altro uovo completo. Questo è un caso piuttosto raro, ma può manifestarsi sia con l'uovo interno che quello esterno che presentano la stessa anomalia. Il caso più frequente, tuttavia, è quello di un uovo completo dentro a un uovo completo. Un caso ancora più raro è un uovo avitellino in un uovo avitellino. Le uova doppie spesso presentano forma anomala, e il guscio esterno è solitamente più grande del normale, mentre quello interno è generalmente più piccolo del normale e ha un guscio sottilissimo.
Altre anomalie possono includere uova con più gusci, uova contenenti parassiti, ghiaia, piume o altri materiali estranei. Le cause di queste anomalie sono diverse e possono essere attribuite a disturbi ormonali, irritazioni dell'ovidutto (come quella descritta anche da Foster), stress o fattori genetici. Ad esempio, il tuorlo può presentarsi sotto forma di banda, che ha un decorso più o meno longitudinale. In alcune uova, la cuticola non è ben presente, ma si trova al di sotto dello strato rugoso, oppure può apparire raggrinzito, più vicina al polo acuto che a quello ottuso. La forma delle uova può essere irregolare, e la presenza di sangue è un'altra anomalia.
Queste variazioni possono essere il risultato di alterazioni nei tempi di ovulazione o nel transito dell'uovo nell'ovidutto. Un ritardo di un'ora e mezza fra due successive ovulazioni, o un uovo che è rimasto nell'ovidutto molto più a lungo del normale, possono portare a uova anomale. Questi eventi possono essere influenzati da fattori come lo stress o la disfunzione ormonale. Ad esempio, disturbi ormonali e la deposizione ritardata causano una maggiore percentuale di anomalie. In particolare, è stata osservata una maggiore incidenza di anomalie nelle galline di razza Plymouth Rock bianca, tenute in gabbie separate.
