La crescita in lunghezza delle ossa è un processo affascinante e finemente regolato, la cui chiave di volta risiede nella cartilagine di coniugazione, nota anche come disco epifisario. Questa struttura, strategicamente posizionata tra la diafisi e l'epifisi, è il fulcro di un'orchestra cellulare e molecolare che determina l'allungamento dello scheletro. Le informazioni riportate di seguito non sono consigli medici e potrebbero non essere accurate.
Struttura e Organizzazione Sequenziale
La cartilagine di coniugazione è caratterizzata da una disposizione sequenziale di cellule cartilaginee ed ossee ordinate regolarmente in colonne che corrispondono ai successivi stadi di ossificazione. Esaminando una sezione longitudinale di un osso lungo, si possono riconoscere, procedendo dal confine con l’epifisi verso la diafisi, altrettante zone che corrispondono ai successivi stadi di ossificazione. Questa organizzazione zonale è fondamentale per comprendere il meccanismo di accrescimento.
Zona della Cartilagine di Riserva o a Riposo
La zona di cartilagine più vicina all’epifisi è formata da cellule cartilaginee distribuite isolatamente o in gruppi isogeni e prive di attività proliferativa. Questa è la regione più distante dalla diafisi, dove i condrociti sono quiescenti e servono come riserva cellulare per le zone sottostanti.

Zona della Cartilagine in Proliferazione
Subito al di sotto della zona di riserva si trova la zona di proliferazione. Qui i condrociti si moltiplicano attivamente formando dei gruppi isogeni rettilinei disposti in colonne parallele all’asse longitudinale dell’abbozzo scheletrico osteo-cartilagineo. Tale disposizione di cellule allineate in colonne è dovuta al fatto che il rigido collare osseo periostale forza la cartilagine a crescere in direzione longitudinale. Le cellule cartilaginee al confine con l’epifisi proliferano attivamente disponendosi in lunghe colonne longitudinali anziché in gruppi isogeni; questa disposizione è resa possibile dal fatto che i solchi di segmentazione delle successive divisioni cellulari sono disposti su piani tra loro paralleli. Le lacune cartilaginee in tale zona, detta zona della cartilagine seriata, sono appiattire con il loro diametro maggiore perpendicolare all’asse longitudinale dell’abbozzo cartilagineo. Questa precisa modalità di proliferazione delle cellule cartilaginee ha un preciso significato morfologico in quanto fa sì che la diafisi si accresca in lunghezza assai più rapidamente che in larghezza. Questa regione assicura il continuo accrescimento in lunghezza del segmento scheletrico.
Istologia - Lezione 10: Cartilagine e Osso pt. 1
Zona della Cartilagine Ipertrofica
Nella zona di ipertrofia o di maturazione delle cellule e di ingrandimento delle lacune, la proliferazione cellulare si arresta ed i condrociti, sempre disposti in colonne parallele, diventano più grandi, ossia ipertrofici, arricchendosi di glicogeno e di lipidi e secernendo fosfatasi alcalina nonché collagene di tipo X, ovvero un marker della ipertrofia condrocitaria. I condrociti ipertrofici secernono anche fattori, come il VEGF, che stimolano l’angiogenesi in questa zona. Le lacune si ingrandiscono notevolmente a spese della matrice interposta. Si mantiene in questa zona la disposizione in colonne longitudinali delle cellule.
Zona della Cartilagine Calcificata
Successivamente, la matrice interposta tra le lacune adiacenti disposte in colonne longitudinali si calcifica, determinando la degenerazione delle cellule, e va incontro a grande regressione riducendosi a sottili setti interposti tra le lacune allargate; in vari punti tali setti scompaiono completamente e si formano ampie cavità longitudinali confluenti. La zona di calcificazione della cartilagine è facilmente riconoscibile per un’accentuata basofilia. Alla regressione della cartilagine contribuiscono forse anche l’azione litica dei condroblasti e quella erosiva dei vasi sanguigni. I condrociti adiacenti alla zona di invasione vascolare non sono affatto in degenerazione, ma sono strutturalmente integri.
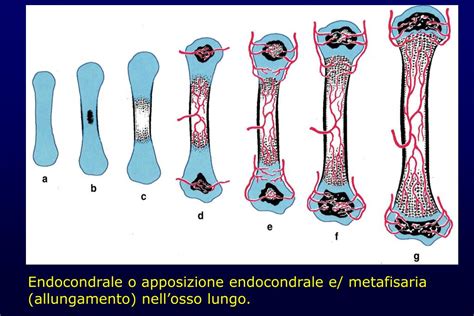
Zona di Invasione Vascolare e di Ossificazione Endocondrale
È la zona dove penetrano i vasi sanguigni, stimolati da segnali proangiogenici condrocitari. I condrociti ipertrofici muoiono per apoptosi e lasciano vuote le lacune; prima di scomparire del tutto i condrociti, a seguito di glicolisi anaerobia, liberano acido lattico che concorre a dissolvere la matrice mineralizzata ampliando le lacune cartilaginee. Condroclasti provenienti dal circolo sanguigno demoliscono le sottili pareti di matrice cartilaginea che separano in senso trasversale le lacune, ampliandone le cavità che andranno a confluire in senso longitudinale. Della ECM originariamente ai lati delle colonne di condrociti residuano solo sottili setti longitudinali di matrice cartilaginea calcificata. Le lacune vengono invase da vasi capillari sanguigni (le cui pulsazioni facilitano i processi demolitivi) e da cellule mesenchimali provenienti dalla cavità midollare diafisaria. Le cavità longitudinali derivanti dalla confluenza delle lacune cartilaginee sono invase da anse capillari e da tessuto connettivo provenienti dagli spazi midollari della diafisi. Nell’ambito di questi tralci connettivali si differenziano cellule emopoietiche e osteoblasti. Dalle cellule mesenchimali osteoprogenitrici si differenziano gli osteoblasti che si insediano sui sopra descritti setti longitudinali di matrice cartilaginea calcificata residua ed iniziano a deporre osteoide. Questi ultimi si dispongono alla superficie delle lamelle cartilaginee in regressione e vi depositano uno strato di tessuto osteoide o preosseo; il tessuto osteoide calcifica a mano a mano che è depositato, diventando tessuto osseo. Le trabecole cartilaginee in regressione svolgono un ruolo importante in quanto circoscrivono cavità che accolgono il midollo osseo primitivo ed i vasi sanguigni e costituiscono una trama di sostegno per la deposizione di sostanza ossea secreta dagli osteoblasti. L’ossificazione endocondrale è il processo in cui le cellule mesenchimali formano prima uno scheletro cartilagineo, che viene poi sostituito da tessuto osseo. I due processi, di accrescimento e di ossificazione, procedono con velocità uguale e come risultato si ha un aumento in lunghezza della diafisi. La cartilagine di coniugazione contribuisce dunque in modo determinante all’accrescimento in lunghezza del segmento scheletrico.
La Regolazione dell'Accrescimento Osseo
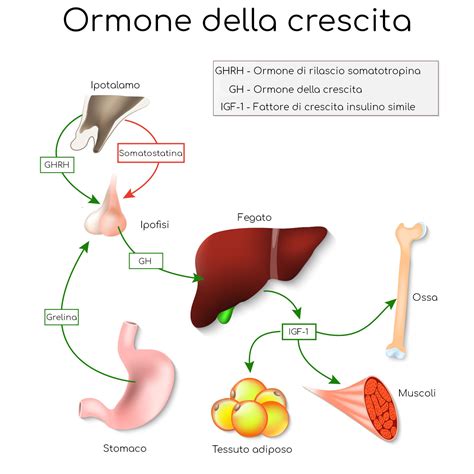
La durata di accrescimento delle ossa lunghe va dal periodo embrionale fino al termine della pubertà. Durante il primo anno di vita, la crescita delle ossa lunghe è veloce, per poi attenuarsi nel decennio successivo. L'attività proliferativa delle cellule della cartilagine di coniugazione è stimolata dall'ormone ipofisario della crescita (GH, growth hormone), il quale induce la produzione di IGF-1 da parte degli epatociti: l’IGF-1, a sua volta, agisce da fattore di crescita sui condrociti. Prima della nascita, si ritiene che IGF-l sia il principale regolatore della crescita più di quanto lo sia il GH. Un adenoma ipofisario durante l'infanzia o l'età adulta provoca una maggiore secrezione di GH, che porta al gigantismo o acromegalia. È interessante notare che diversi studi hanno dimostrato che l'estrogeno può inibire la crescita longitudinale in assenza di GH. La famiglia delle BMPs (Bone Morphogenetic Proteins) è composta da almeno 15 membri, che fanno tutti parte della famiglia del TGFß. Questi fattori di crescita e differenziazione giocano un ruolo cruciale nella formazione e riparazione del tessuto osseo, influenzando anche l'attività della cartilagine di coniugazione.
Fratture della Cartilagine di Coniugazione
La cartilagine di coniugazione, fino a che non è completamente chiusa, costituisce la parte più fragile dell'osso e sono perciò frequenti fratture e lesioni, che possono addirittura estendersi nell’epifisi o nella diafisi. Queste lesioni, particolarmente comuni in età pediatrica, sono classificate secondo diversi tipi. Nei tipi da I a IV la cartilagine di coniugazione è separata dalla metafisi. Un esempio è la frattura di Salter di livello II, una frattura a scheggia della cartilagine di coniugazione, che può essere molto piccola. Un altro tipo è la frattura di Salter di livello V, una frattura da compressione della cartilagine di coniugazione. Spesso, le fratture della cartilagine di coniugazione sono accompagnate da dolore e gonfiore localizzato nella zona lesa e da una difficoltà o addirittura impossibilità nel muovere l'arto fratturato. La gestione di queste fratture è cruciale per prevenire alterazioni della crescita e deformità ossee.
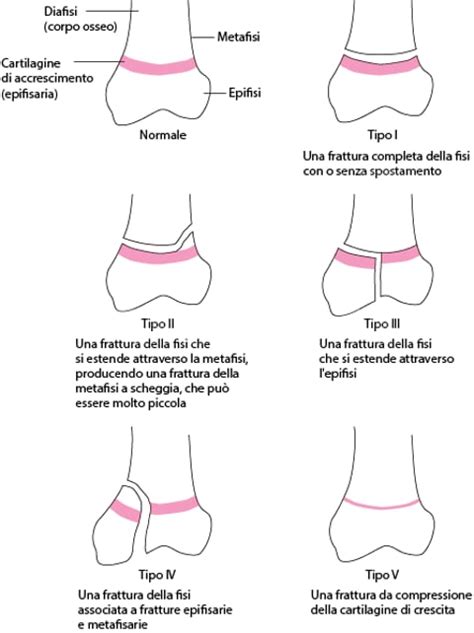
La Chiusura delle Epifisi
Al termine del periodo di accrescimento, l’attività proliferativa dei condrociti rallenta e poi si arresta mentre il processo di ossificazione procede fino alla sostituzione completa della cartilagine di coniugazione con tessuto osseo; tale processo è denominato chiusura delle epifisi. La zona di confine tra epifisi e diafisi è visibile nell’adulto come una linea sottile, detta linea epifisaria. Da questo momento non è più possibile un ulteriore aumento in lunghezza del segmento scheletrico.
Istologia - Lezione 10: Cartilagine e Osso pt. 1
Approfondimenti sui Tessuti Coinvolti
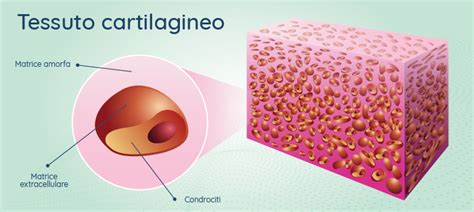
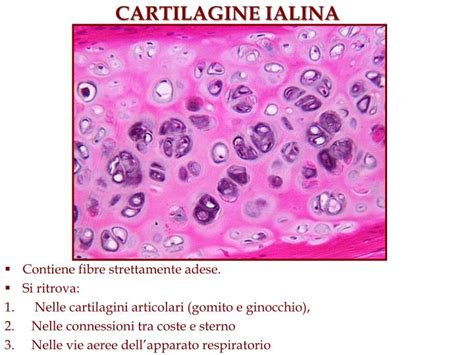
Il tessuto cartilagineo è un tessuto connettivo specializzato, costituito da cellule, i condroblasti e i condrociti, circondate da una matrice extracellulare allo stato di gel con fibre. La cartilagine è l’unico tessuto connettivo sprovvisto di vasi sanguigni, per cui il nutrimento di questo tessuto è affidato alla permeabilità della matrice extracellulare. La cartilagine è generalmente rivestita da un connettivo capsulare, il pericondrio, fatta eccezione per le articolazioni dove non presenta pericondrio ed è a diretto contatto con il liquido sinoviale. È sicuramente il tipo di cartilagine più rappresentato nei mammiferi. I condrociti presentano un nucleo ben evidente con uno o più nucleoli e sono alloggiati in lacune o depressioni della matrice extracellulare. Quando la cartilagine ialina è matura, i condrociti tendono a raggrupparsi in isolotti, i gruppi isogeni. Questi sono più numerosi nella zona profonda della cartilagine e più scarsi nelle zone più periferiche (zona intermedia e zona tangenziale). I gruppi isogeni sono generalmente assenti nella cartilagine ialina immatura. La cartilagine ialina è avascolare e generalmente circondata da pericondrio. La matrice, povera di fibre, è formata prevalentemente da sostanza amorfa ricca di proteoglicani.
La cartilagine fibrosa, presente in alcune cartilagini articolari come i dischi intervertebrali e i menischi del ginocchio, rappresenta una forma di transizione tra la cartilagine ialina ed il tessuto connettivo denso, tanto che non è possibile avere un confine netto fra i due. Si presenta sotto forma di piccole placche a confini poco distinti, costituite da condrociti, da una scarsa quantità di sostanza fondamentale e da abbondanti elementi fibrosi (collagene di tipo I). È generalmente avascolare e non presenta pericondrio. I condrociti, circondati da matrice extracellulare, sono isolati, in coppia o, talvolta, allineati gli uni sugli altri a formare una lunga fila tra i fasci di fibre collagene. In un disco intervertebrale umano, nella zona di transizione tra cartilagine ialina e cartilagine fibrosa, la matrice extracellulare della cartilagine ialina assume un aspetto fibroso e perde gradatamente la sua acidità e, di conseguenza, anche il colore viola dell'emallume. Nella cartilagine fibrosa i condrociti tendono a diminuire, sia nel numero che nelle dimensioni. Le fibre collagene si allungano, si inspessiscono e vanno così a formare la cartilagine fibrosa. Le cellule cartilaginee sono alloggiate nelle lacune disposte in file parallele e separate da denso tessuto collagene. Più il tessuto tende ad essere fibroso e più la colorazione passa dal viola, tipico della cartilagine, al rosso-marrone tipico del collagene.
Il tessuto osseo è un tessuto dinamico e plastico: provvede a modulare la propria struttura in seguito a stimoli organici e meccanici. Esso è formato da una parte organica ed una parte inorganica. La parte organica è composta da cellule proprie del tessuto osseo (cellule osteoprogenitrici, osteoblasti, osteociti e osteoclasti che provvedono rispettivamente alla formazione, all’accrescimento, alla produzione, e al riassorbimento del tessuto osseo) e da matrice extracellulare (sostanza amorfa e fibre collagene di tipo I). La parte inorganica della matrice extracellulare è costituita da numerosi sali minerali come i fosfati di calcio e magnesio e i citrati di Na, Mn, K. La componente organica della matrice extracellulare rappresenta il 35% del peso secco dell’osso e ne determina robustezza ed elasticità mentre la componente inorganica mineralizzata rappresenta il 65% del peso secco e conferisce al tessuto osseo compattezza e durezza.
Le cellule osteoprogenitrici sono cellule di origine mesenchimale con proprietà staminali: esse possono proliferare e differenziare in osteoblasti. Gli osteoblasti sono i precursori degli osteociti; sono cellule voluminose, altamente polarizzate, con un nucleo ovoidale leggermente spostato in periferia e con citoplasma intensamente basofilo. Gli osteoblasti provvedono alla produzione sia della matrice organica (definita osteoide) che alla deposizione di quella inorganica, hanno quindi funzioni osteogeniche. Quando gli osteoblasti hanno terminato la formazione dell’osso rimanendo intrappolati all’interno di lacune nella matrice da loro stessi prodotta, diventano osteociti. Gli osteociti sono cellule di forma irregolare, con un nucleo ben evidente e un citoplasma che presenta diversi prolungamenti. Essi sono alloggiati nelle lacune ossee dalle quali si dipartono, in ogni direzione, numerosi canalicoli microscopici. Attraverso questi canalicoli i prolungamenti citoplasmatici di osteociti diversi prendono contatto tra di loro tramite giunzioni comunicanti e con capillari sanguigni presenti nei canali ossei, permettendo così scambi metabolici tra gli osteociti stessi e tra osteociti e sangue. Gli osteoclasti non appartengono alla linea osteoprogenitrice ma derivano dalla fusione di numerosi precursori monocitari (fino a 30) e sono deputati alla distruzione (riassorbimento) e al rimaneggiamento del tessuto osseo. Sono cellule molto grandi, potendo superare anche i 100 µm di diametro, e hanno numerosi nuclei. Anche gli osteoclasti sono cellule altamente polarizzate: quando attivate presentano una faccia citoplasmatica in prossimità dell’osso con caratteristiche increspature molto mobili ed aderiscono alla superficie dell’osso creando un microambiente isolato da quello circostante (zona sigillata) che viene acidificato per l’attivazione successiva di enzimi di derivazione sia lisosomale (proteinasi e fosfatasi) che non lisosomale (metalloproteinasi).
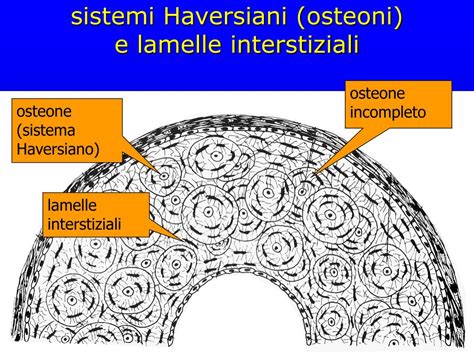
Nel tessuto osseo non lamellare a fibre intrecciate le fibre collagene sono intrecciate a formare un fitto reticolo. La sostanza fondamentale, disposta irregolarmente, è poco rappresentata sia nella sua parte organica che inorganica. Le lacune ossee hanno forma globosa e sono tendenzialmente più grandi che nel tessuto osseo lamellare. Il tessuto osseo lamellare, grazie alla sua composizione chimica e alla sua particolare organizzazione strutturale, ha una forte resistenza alla trazione, alla pressione e alle sollecitazioni meccaniche in generale. Nelle ossa lunghe si distinguono le epifisi, corte e rotondeggianti, localizzate alle estremità e costituite prevalentemente da osso spugnoso, e una parte allungata, centrale, a forma di cilindro cavo detta diafisi, formata da osso compatto, estremamente resistente agli urti e alle pressioni e contenente midollo osseo. Nelle ossa piatte, invece, distinguiamo due superfici di tessuto osseo compatto, denominate tavolato interno ed esterno. Il tessuto osseo compatto costituisce le diafisi nelle ossa lunghe, lo strato superficiale delle epifisi, i tavolati delle ossa piatte e in generale riveste tutte le superfici ossee. Le lamelle ossee sono organizzate in strati concentrici a formare gli osteoni in cui gli osteociti sono posti circolarmente, secondo diverse orbite, intorno a canali, detti canali di Havers, contenenti uno o due piccoli capillari sanguigni. Il numero di lamelle che circonda un singolo canale di Havers può variare da 4 a 20. I vari canali di Havers comunicano tra di loro grazie ad altri canali posti, però, trasversalmente od obliquamente, detti canali di Volkmann, garantendo un'efficiente irrorazione sanguigna e scambio di nutrienti all'interno del tessuto osseo compatto.
tags: #cartilagine #fertile #verso #epifisi