Lo sviluppo umano, un processo di incredibile complessità e delicatezza, ha inizio con l'unione di due cellule specializzate: l'ovocita e lo spermatozoo. I fenomeni biologici che portano alla formazione di un embrione sono innumerevoli e soprattutto estremamente delicati e complessi. Questo percorso, che parte dalla formazione dello zigote, la prima cellula che si forma dell’embrione e quindi la prima cellula del nuovo individuo, attraversa diverse fasi di trasformazione e differenziazione cellulare, culminando nella formazione della blastocisti e nello sviluppo degli annessi embrionali, strutture indispensabili per il completamento dello sviluppo. La coltura embrionaria può quindi concludersi in fasi differenti che possono andare dal giorno 2 o 3 fino al giorno 5 - 6 - 7.
Il Miracolo del Concepimento: Dalla Gametogenesi allo Zigote
Il punto di partenza di ogni nuovo individuo è la gametogenesi, il processo di sviluppo dalle cellule germinali primordiali ai gameti maturi. Nelle femmine, questo processo è chiamato oogenesi, mentre nei maschi è noto come spermatogenesi. Nelle femmine e nei maschi, inizia con cellule germinali diploidi che poi vanno incontro a mitosi, meiosi e citodifferenziazione in gameti aploidi.
La formazione delle cellule uovo immature (oogoni) avviene già nella vita intrauterina. Fin dall’infanzia di una donna si sviluppano i follicoli nei quali si trova l’ovocita, ma a partire dalla pubertà solo uno o, occasionalmente, due di essi raggiungeranno la maturità ogni mese durante il ciclo mestruale.
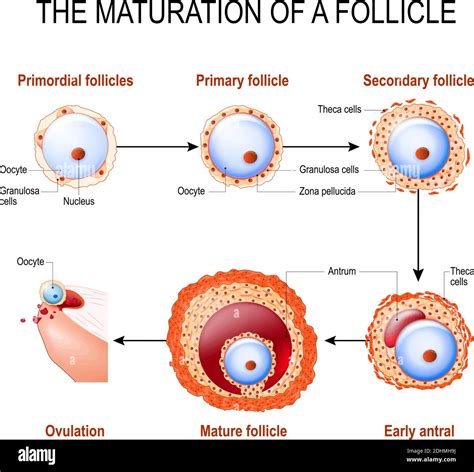
Nel frattempo, nei testicoli maschili possono essere prodotti miliardi di spermatozoi. L'ovulazione si verifica circa 14 giorni dopo il primo giorno di un ciclo mestruale in una donna con cicli mestruali di 28 giorni. Al momento dell'ovulazione, il muco cervicale diventa meno viscoso, facilitando il rapido movimento degli spermatozoi verso l'ovocita. Lo spermatozoo può sopravvivere nelle vie genitali fino a circa 3 giorni dopo il rapporto sessuale.
L'incontro reale tra spermatozoo e cellula uovo si ha durante la fecondazione (concepimento), che avviene, nella quasi totalità dei casi, nell'ampolla delle tube uterine. Gli spermatozoi (gameti aploidi maschili) migrano attraverso il canale cervicale, la cavità uterina e nelle tube di Falloppio. L'ovocita entra nella tuba di Falloppio attraverso l'estremità fimbriata, viaggia attraverso la tuba di Falloppio e poi passa nella cavità uterina. La fecondazione si verifica in genere quando l'ovocita si trova nella tuba di Falloppio. Nella specie umana la fecondazione si ha per monospermia, ovvero un solo spermatozoo entra nella cellula uovo e fonde il proprio nucleo con quello della cellula uovo matura. Di questi miliardi di spermatozoi, solo uno riuscirà, dopo l’eiaculazione, ad attraversare la corona radiata che circonda l’ovulo e a superare la membrana.
L'ovocita diventa un ovulo fecondato quando uno spermatozoo penetra negli strati esterni dell'ovocita. In genere, 1 spermatozoo fertilizza 1 ovocita. L'ovulo fecondato, lo zigote, è diploide. Questo si forma grazie all’unione del DNA dello spermatozoo (23 cromosomi) con quello dell’ovocita (23 cromosomi). Tuttavia, 2 ovociti possono essere rilasciati e poi fecondati da 2 spermatozoi, con conseguente gravidanza gemellare dizigotica (fraterna). È importante notare che, in alcune rare circostanze, può essere rilasciato più di un ovulo durante un ciclo mestruale. Quando ciò accade e più di uno di questi ovuli viene fecondato con successo da spermatozoi diversi, può verificarsi una gravidanza gemellare. Una gestazione multipla di ordine superiore (p. es., triplette, quadruplette) può verificarsi se vengono liberati e fecondati più di 2 ovociti, ma questo è raro. Inoltre, più di 1 spermatozoo può penetrare un singolo ovocita, ma questo si traduce in uno zigote anormale.
Le Prime Fasi dello Sviluppo Embrionale: Dallo Zigote alla Morula
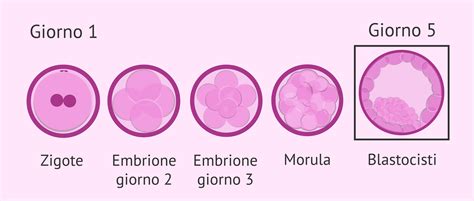
Dopo la formazione dello zigote, il processo di sviluppo embrionale procede attraverso diverse fasi embrionali prima dell’annidamento nell’endometrio. Lo zigote si divide ripetutamente mentre migra verso il sito di impianto nell'endometrio (di solito vicino al fondo o sulla parete posteriore dell'utero). Per i primi tre o quattro giorni dopo la fecondazione, lo zigote scende lungo le tube di Falloppio assorbendo sostanze nutritive dalle secrezioni materne e andando incontro alle prime divisioni mitotiche. Le cellule figlie che derivano dalle prime divisioni dello zigote sono chiamate blastomeri. All’inizio sono totipotenti, cioè ciascuna può teoricamente dare origine a un intero organismo con i suoi annessi.
La coltura embrionaria può quindi concludersi in fasi differenti che possono andare dal giorno 2 o 3 fino al giorno 5 - 6 - 7. Nella seconda giornata dalla fecondazione, gli embrioni possono aver raggiunto lo stadio di due cellule chiamate blastomeri. Successive divisioni portano l’embrione allo stadio di sei/otto blastomeri nella terza giornata. Nel passaggio da 8 a 16 cellule, l’embrione inizia un processo detto di “compattazione”. Le cellule (chiamate blastomeri) si schiacciano le une sulle altre formando una massa cellulare compatta, che massimizza i contatti tra una cellula e le circostanti. La compattazione e’ un evento fondamentale per questa trasformazione. Le cellule che lo compongono, chiamate blastomeri, iniziano ad interagire tra di loro formando dei canali che permettono loro di avvicinarsi e compattarsi formando la morula.
A questo stadio l’embrione si trova al 4° giorno del suo sviluppo e viene chiamato morula. La morula continua ad aumentare esponenzialmente il numero delle sue cellule. Diversamente dallo zigote, che è un embrione composto da un massimo di sedici cellule, la morula si caratterizza per una quantità di cellule variabile tra sedici e sessantaquattro. Questa palla compatta di cellule ha un aspetto simile a una mora.
La Struttura Complessa della Blastocisti: Specializzazione Cellulare e Formazione del Blastocele
A partire dalla fine della terza giornata, l’embrione comincia a subire una profonda trasformazione della propria struttura. La compattazione precede la blastulazione. Quando la morula continua a dividersi, si trasforma gradualmente in una “blastocisti” (circa cinque-sei giorni dopo la fecondazione).
All’interno della morula, tra la quinta e la settima giornata, comincia a formarsi uno spazio composto di liquido che mano a mano si espande. Questa cavità è detta blastocele. L’accumulo di fluido all’interno della massa cellulare - una cavità, detta blastocele, si espande progressivamente. Questa fase è caratterizzata dalla formazione di una cavità centrale chiamata “blastocele”. Il blastocele spinge le cellule verso l’esterno, a contatto con la zona pellucida. Si forma quindi la blastocisti.
Al momento dell'impianto, lo zigote è divenuto una blastocisti, che è uno strato di cellule che circonda una cavità. La parete della blastocisti ha un singolo strato di cellule eccetto il polo embrionale, formato da 3 o 4 strati di cellule. La blastocisti è costituita da una parete di cellule appiattite (il trofoectoderma, da cui si formeranno gli annessi embrionali) e da un gruppo di cellule (la massa cellulare interna, dalle quali si formerà il bottone embrionario). In seguito alla segmentazione, l'embrione si manifesta con la forma di una sfera cava che prende il nome di blastocisti. Nella blastocisti possiamo individuare due parti principali: il “trofoblasto” (o “trophectoderma”), che diventerà la placenta, e l’embrione stesso. La blastocisti presenta una cavità piena di liquido, una massa interna di cellule da cui si formerà il feto e uno stato esterno di cellule detto trofoblasto (o sincizio-trofoblasto). L’architettura dell’embrione a questo punto è completamente cambiata.
Con la compattazione e la cavitazione, le cellule iniziano a specializzarsi: quelli più esterni formano il trofoectoderma, quelli interni costituiscono la massa cellulare interna. Possiamo definire l’embrione come la fase iniziale dello sviluppo di un organismo multicellulare che segue il momento del concepimento. Nel corso di un trattamento di fecondazione in vitro, l’ovocita viene inseminato in laboratorio e l’embrione risultante viene conservato e monitorato per un periodo variabile dai due ai sei giorni, durante i quali si sviluppa in un organismo complesso pronto all’impianto nell’utero. Durante questo periodo, l’embrione subisce divisioni cellulari continue seguendo un ritmo specifico. Tra la centesima e la centoventesima ora dalla fecondazione si verifica invece la blastulazione.
L'Annidamento: Il Momento Critico dell'Impianto Uterino
Verso la fine della prima settimana, la blastocisti si annida nell'utero e va ad aderire all'endometrio. Il momento in cui l’embrione viene trasferito nell’utero varia infatti da caso a caso. La blastocisti ha inoltre peculiarità strutturali del tutto diverse dall’embrione dei giorni precedenti e si compone di due aree che ne indicano la qualità e la possibilità di impianto e gravidanza. Come è facile intuire, lo stadio di blastocisti è molto importante per la successiva fase di impianto nell’utero materno.
La blastocisti è rotolata nella cavità dell’utero grazie all’aiuto delle ciglia che l’hanno mossa fino a farla posizionare con la massa cellulare interna appoggiata sulla mucosa dell’utero. A questo punto inizia la fase di annidamento e la blastocisti impianta le sue radici nella mucosa dell’utero, ancorandosi in modo tale da non potersi più spostare. Circa 6 giorni dopo la fecondazione, la blastocisti si impianta nell'endometrio; il polo embrionale, che si svilupperà nell'embrione, è il primo punto dell'impianto. L'annidamento o impianto è la terza e ultima fase dello sviluppo pre-embrionale nei mammiferi placentati, ossia del periodo comprendente le prime due settimane di sviluppo.
Il trofoblasto secerne enzimi litici che permettono l'impianto della blastocisti nell'endometrio, lo strato cellulare che riveste internamente la cavità uterina. Questi enzimi litici hanno una funzione digestiva e permettono alla blastocisti di annidarsi; a contatto con la mucosa digeriscono la mucosa uterina e piano piano si scavano una specie di nicchia nella quale viene accolta la blastocisti. Il trofoblasto è costituito da estroflessioni costituite da cellule in attiva divisione che penetrano nell'endometrio, delle radici di annidamento, e mano a mano che si formano scendono un po' a caso nella mucosa uterina. Alcune si fondono tra loro generando delle nicchie contenenti tonaca propria; queste porzioni di tonaca propria vengono aggredite dagli enzimi, digerite (quindi rese liquide) e poi assorbite dal sincizio stesso e utilizzate come nutrimento.
Durante la segmentazione e la gastrulazione l'embrione in sviluppo si nutre dalla secrezione delle ghiandole uterine o dei condotti uterini. Queste ghiandole producono un secreto (il cosiddetto "latte uterino") molto ricco di glicogeno. L'annidamento viene completato con l'erosione della parete dei vasi sanguigni materni, circa 14 giorni dopo la segmentazione. Quando i vasi sanguigni della madre vengono rotti, il sangue materno invade, con una certa pressione, le cosiddette lacune del sinciziotrofoblasto, cioè gli spazi vuoti lasciati dal tessuto digerito. Queste lacune comunicano un po' tutte tra di loro. Poiché il sangue penetra con una certa pressione, passando tra una lacuna e l'altra e intorno al sincizio, può addirittura rompere il tappo di fibrina che si è formato nel punto di annidamento. Di conseguenza può succedere che si abbia una perdita di sangue nella cavità dell'utero, che, tramite le vie genitali femminili, esca all'esterno.
L'annidamento, nella maggioranza dei casi, avviene sul fondo dell'utero (la parte più alta di quest'organo), anche se ci sono dei casi in cui avviene prima. Quando la blastocisti è ancora nelle tube uterine, l'annidamento avviene ugualmente perché la struttura delle tube uterine è praticamente uguale a quella dell'utero; questo è un caso di gravidanza extrauterina. Le prime fasi procedono senza complicazioni, però non è previsto che una tuba uterina, per quanto elastica possa essere, possa contenere né un feto né tanto meno un neonato. Il rischio che si corre è che ad un certo punto esse si rompano e che, essendo organi estremamente vascolarizzati, si possa avere una rischiosa emorragia. Proprio per questo motivo solitamente le gravidanze extrauterine vengono interrotte; una gravidanza di questo tipo può andare avanti solo se l'impianto di questa blastocisti è situato proprio allo sbocco della tuba uterina nell'utero.
LA FECONDAZIONE UMANA
Intorno al 7° giorno dopo la fecondazione, che corrisponde ai giorni immediatamente successivi al transfer in un percorso di FIV, l’embrione si posiziona sull’endometrio, individuando il sito ideale per l’annidamento. In questa fase, i pinopodi-protrusioni endometriali che appaiono solo durante la finestra d’impianto-guidano l’embrione verso l’endometrio, facilitando l’interazione iniziale. Successivamente, l’embrione penetra l’endometrio. Le cellule epiteliali endometriali si riorganizzano per agevolare l’invasione embrionale nel tessuto stromale endometriale. A questo punto, l’embrione stabilisce contatti con i vasi sanguigni materni, processo cruciale per garantirne il nutrimento e lo sviluppo.
La finestra di impianto rappresenta il periodo in cui l’endometrio è recettivo per l’annidamento embrionale. Il progesterone, prodotto dal corpo luteo nell’ovaio dopo l’ovulazione, aumenta lo spessore dell’endometrio e favorisce la produzione di sostanze chimiche che consentono all’embrione di aderire. La Gonadotropina Corionica Umana (hCG) è invece un ormone, prodotto dall’embrione dopo l’annidamento, che segnala al corpo luteo di continuare a produrre progesterone. Il mantenimento del progesterone infatti è essenziale per evitare il rifiuto dell’embrione da parte del sistema immunitario materno. Le selectine e le molecole di adesione svolgono un ruolo cruciale nell’attaccamento dell’embrione all’endometrio. Nel contesto della fecondazione in vitro, l’endometrio viene preparato farmacologicamente per simulare questa finestra d’impianto. Negli ultimi anni sono stati sviluppati test specifici, come l’Endometrial Receptivity Analysis (ERA), per aiutare i medici a individuare la finestra d’impianto.
Gli Annessi Embrionali: Supporto e Protezione per l'Embrione in Sviluppo
Gli annessi embrionali sono strutture indispensabili per il completamento dello sviluppo embrionale del germe ma che, una volta arrivati al momento della nascita, vengono abbandonati poiché non servono più. Sono detti anche membrane extraembrioniali, poiché proteggono l'embrione.
Il primo annesso embrionale che si forma è l'amnios, che si forma per cavitazione. Alcune cellule della massa cellulare interna vanno incontro ad apoptosi, cioè morte cellulare programmata. Si forma una cavità, in cui si distinguono una volta e un pavimento, chiamata cavità amniotica, ripiena di liquido secreto dagli amniociti. La volta rimane aderente al citotrofoblasto mentre il pavimento è formato da due strati di cellule: lo strato immediatamente a contatto con la cavità amniotica è il primo foglietto embrionale ed è una formazione dell'ectoderma, invece lo strato sottostante è dell'endoderma.
Nella cavità amniotica è presente un liquido, il cosiddetto liquido amniotico, che ha la funzione di attutire gli urti e di impedire la disidratazione del feto. Il liquido è incomprimibile, perciò una pressione che si esercita su un punto viene suddivisa su tutta la superficie, in modo tale che l'urto risulti minore. L'amnios all'inizio è situato al di sopra del germe, quando il germe è ancora allo stato bilaminare. Poi la vescicola amniotica si amplia, cresce in tutti i sensi piuttosto velocemente e va a delimitare la forma dell'embrione che dapprima diviene allungata in senso antero-posteriore e non è più laminare. Continuando a svilupparsi, la vescicola amniotica che è cresciuta in tutti i sensi intorno all'embrione, si stringe poi ventralmente sotto l'embrione in corrispondenza di quello che diventerà l'ombelico.
Dalla parte opposta dell'amnios, dal cito-trofoblasto si differenzia un altro gruppo di cellule che danno luogo ad una lamina monostratificata, la quale comincia a moltiplicarsi rimanendo anch'essa su un unico strato e scivolando sul cito-trofoblasto. Queste due lamine si toccano, si vanno a fondere l'una nell'altra e si forma il sacco vitellino primario, la cui parete, quindi, è formata dall'endoderma e da questa membrana, che deriva dal citotrofoblasto. Sempre dal cito-trofoblasto si forma un altro strato di tessuto che si differenzia tra la membrana che si è appena formata e il citotrofoblasto, e le cellule di questo tessuto iniziano a moltiplicarsi e ad espandersi scivolando tra i due strati. Questo strato continua a crescere finché le due estremità non si saldano tra loro.
Nella parte inferiore, il nuovo tessuto, il mesoderma extraembrionale (si trova al di fuori dell'embrione) va ad interporsi tra sacco vitellino primario, citotrofoblasto e sinciziotrofoblasto nella parte inferiore, e tra amnios e citotrofoblasto nella parte superiore. Questo mesoderma cresce molto velocemente, soprattutto nella parte intorno al sacco vitellino primario; quest'ultimo viene strozzato dalla crescita del mesoderma extraembrionale che cresce principalmente in corrispondenza del punto di contatto tra endoderma e la lamina monostratificata (quella con cui l'endoderma è andato a fondersi). Il sacco vitellino primario viene trasformato in una specie di clessidra, finché ad un certo punto viene staccata la membrana (lamina monostratificata) che si stacca perché le estremità dell'endoderma si fondono tra loro. Questa membrana viene staccata e poi scompare e l'endoderma si salda su sé stesso.
Dentro il mesoderma extraembrionale cominciano a formarsi, per cavitazione (come per l'amnios), delle cavità piene di liquido le quali confluiscono l'una nell'altra fino a formare un'unica grande cavità. Quindi questo mesoderma extraembrionale va a costituire due lamine, la lamina parietale, quella più esterna che è accollata sempre al cito-trofoblasto, e la lamina viscerale, quella più interna, che va a costituire il sacco vitellino secondario accollandosi all'endoderma del sacco vitellino primario. La cavità rimane la stessa perché la parete del sacco vitellino primario è formata da endoderma e dalla membrana che si era formata dal citotrofoblasto; poi questa membrana viene staccata e riassorbita, le estremità dell'endoderma si saldano tra loro e all'endoderma si accolla il foglietto viscerale che si è formato dal mesoderma extraembrionale. Quando la parete non è più formata da endoderma e membrana, bensì da endoderma e foglietto viscerale del mesoderma extraembrionale, è definito sacco vitellino secondario. La differenza non risiede nella cavità, bensì nelle lamine che formano la parete: la parete del sacco vitellino primario è formata da un solo foglietto, che è l'unione tra endoderma e membrana, quindi è un unico strato di cellule. Dalla parete del sacco vitellino secondario si forma la maggior parte dei vasi sanguigni dell'embrione, i vasi vitellini. Mano a mano che si sviluppa, il sacco vitellino secondario viene un po' risucchiato all'interno dell'embrione e va a costituire una parte interna dell'embrione e una parte va, insieme all'allantoide, a costituire una parte del cordone ombelicale. La parete del sacco vitellino secondario è formata da un doppio strato di cellule, uno interno che è l'endoderma e uno esterno che è il foglietto viscerale e che deriva dalla proliferazione e dalla delaminazione del mesoderma extraembrionale. Una parte del foglietto viscerale si accolla all'endoderma e alla parete del sacco vitellino secondario, mentre l'altra parte si accolla alla parete dell'amnios.
Entro 1 o 2 giorni dall'impianto, uno strato di cellule (cellule del trofoblasto) si sviluppa intorno alla blastocisti. Lo strato interno (amnios) e uno strato esterno (corion) delle membrane si sviluppano a partire dal trofoblasto; queste membrane formano il sacco amniotico, che contiene il prodotto del concepimento. Le cellule del trofoblasto si differenziano in cellule che formano la placenta. Il trofoblasto extravilloso forma i villi, che penetrano nell'utero. Il sinciziotrofoblasto copre i villi. Il sinciziotrofoblasto sintetizza ormoni trofici e fornisce scambi arteriosi e venosi tra la circolazione embrionale e quella materna. Dalla formazione del corion deriva la parte fetale della placenta, che costituisce l'inizio della formazione di essa. Alla formazione del corion partecipano anche il cito-trofoblasto e il sincizio-trofoblasto. La placenta è completamente formata entro le 18-20 settimane ma continua a crescere, fino a raggiungere il peso di circa 500 g al termine della gravidanza.
La delaminazione del mesoderma extraembrionale non è ancora completa: si forma una grossa cavità, anche se nella parte posteriore del germe rimane "indelaminato". Questa parte di mesoderma extraembrionale "non delaminata" si chiama peduncolo d'attacco, e non si delamina perché altrimenti il germe rimarrebbe galleggiante all'interno, senza contatti con la mucosa uterina. Dalla parte posteriore del sacco vitellino secondario, vicino alla giunzione amnio-ectodermica, si forma un'evaginazione, una sporgenza di questo ectoderma che diventa sempre più profonda ed entra dentro il peduncolo d'attacco. Questa si svilupperà notevolmente e formerà l'allantoide, un'evaginazione della parete posteriore della parte del sacco vitellino secondario che entra nel peduncolo d'attacco. Nell'interno del magma reticolato (reticolo extraembrionale) del peduncolo d'attacco che si sta trasformando per diventare il cordone ombelicale, si trova l'allantoide e il residuo del sacco vitellino secondario, ovvero quella parte che non è stata risucchiata dall'embrione. Il peduncolo d'attacco, l'unica parte non delimitata dal mesoderma extraembrionale, che all'inizio, nel germe, era posteriore, dopo che l'amnios si è espanso e ha delimitato le forme dell'embrione, va a ritrovarsi ventralmente in corrispondenza anch'esso dell'ombelico.
Gastrulazione e Organogenesi Iniziale: La Definizione del Piano Corporeo
Quando il sacco è formato e la cavità della blastocisti si chiude (approssimativamente entro il 10o giorno), il prodotto del concepimento viene considerato un embrione. Durante la seconda settimana di gravidanza inizia la gastrulazione, che si conclude con la formazione dei tre foglietti embrionali. Intorno al 10o giorno circa dopo la fecondazione si possono solitamente distinguere all'interno dell'embrione 3 foglietti germinativi (ectoderma, mesoderma, endoderma). Questi foglietti germinativi sono le prime tre lamine di tessuto che si formano e si differenziano, e dalle quali si differenzieranno tutti i tessuti del nuovo individuo.
Osservato dall'alto si vede come un disco, il cosiddetto disco embrionale, che corrisponde al pavimento dell'amnios. A questo stadio di sviluppo, il disco embrionale è circondato dall'amnios e dal corion, tranne nel punto dove un peduncolo lo unisce alla parete interna del corion. Nel disco embrionale compaiono alcune formazioni, come ad esempio la linea primitiva, una linea sagittale e, contemporaneamente, il germe, che da circolare diventa allungato. Questa linea primitiva è una specie di solco lungo il quale una serie di cellule dell'ectoderma sprofondano e si posizionano tra ectoderma ed endoderma. Attraverso divisioni e migrazioni cellulari si forma sul disco embrionale una depressione, detta stria primitiva, che segnala il punto di inizio della gastrulazione e definisce anche l'asse antero-posteriore dell'embrione e, successivamente, la sua simmetria bilaterale. A questo livello, le cellule che migrano verso l'interno danno origine all'endoderma e al mesoderma.
L'ectoderma, considerato il primo foglietto germinativo anche se si forma insieme all'endoderma, è il foglietto più esterno. Da esso derivano tutti gli epiteli di rivestimento composti (o stratificati) come ad esempio l'epidermide e la mucosa del cavo orale. Dall'endoderma derivano tutti gli epiteli semplici (o monostratificati), tra cui l'epitelio di rivestimento interno del tubo digerente. Le cellule dell'ectoderma scendono lungo la linea primitiva e poi si allargano tra lo strato dell'ectoderma e lo strato dell'endoderma e si espandono in tutti i sensi. In avanti, poi si separano e formano le cosiddette ali mesodermiche che, nella parte anteriore dell'embrione, si risaldano tra loro isolando una zona centrale senza mesoderma. Quest'area inizialmente priva di mesoderma è la placca precordale, il punto in cui inizia a formarsi la corda dorsale che poi sarà sostituita dalla colonna vertebrale. È il punto di partenza dell'asse del nostro organismo.
Poi inizia a svilupparsi la stria primitiva, che diventa poi il tubo neurale. Per quanto riguarda la formazione del sistema nervoso, nell'ectoderma si forma un solco longitudinale, analogamente alla formazione della linea primitiva, dalla testa alla coda. Ai lati di questo solco l'ectoderma si solleva formando due creste che poi si saldano tra loro e vanno a costituire una specie di tubo, un canalicolo che è l'abbozzo del sistema nervoso centrale, il tubo neurale, inizialmente aperto sia anteriormente sia posteriormente, per mezzo del neuroporto anteriore posteriore, i quali successivamente si chiudono. Longitudinalmente, c'è una vescicola cerebrale che è l'abbozzo dell'encefalo, e il resto che è l'abbozzo del midollo spinale.
Nel punto in cui le ali mesodermiche si saldano, quindi nel margine anteriore dell'embrione, è situata l'area cardiogenica, il punto in cui si avrà l'abbozzo del cuore. Intorno al 16o giorno la porzione cefalica del mesoderma si ispessisce formando un canale centrale che darà origine al cuore e ai grossi vasi. Il cuore inizia a pompare il plasma intorno al 20o giorno e il giorno successivo compaiono i globuli rossi fetali, che sono immaturi e nucleati. I globuli rossi fetali sono subito sostituiti da globuli rossi maturi e i vasi sanguigni si sviluppano in tutto l'embrione. Infine, si formano l'arteria e la vena ombelicale che collegano i vasi dell'embrione alla placenta.
L'Evoluzione da Embrione a Feto: Una Crescita Rapida e Delicata
La maggior parte degli organi si forma tra i 21 e i 57 giorni dopo la fecondazione (5-10 settimane di gestazione); tuttavia, il sistema nervoso centrale continua a svilupparsi durante tutta la gravidanza. Quando il sacco amniotico si riempie di liquido e si espande al crescere dell'embrione, riempie la cavità endometriale verso le 12 settimane circa dopo il concepimento; a questo punto, il sacco amniotico è la sola cavità rimanente nell'utero. L'embrione misura 4,2 cm.
Molte evidenze e ricerche, anche recenti, hanno dimostrato come le prime 9 settimane di gravidanza rappresentino una fase delicatissima dello sviluppo embrionale. In particolare, l’ambiente uterino diventa responsabile del corretto differenziamento e sviluppo dei tessuti e degli organi e del loro funzionamento. L'evoluzione del mesoderma embrionale è legata strettamente all'evoluzione dell'amnios. Grazie all’evoluzione dell’amnios, il germe passa alla fase tridimensionale e prima assume una forma allungata a bastoncino; successivamente si formano la curva cefalica e la curva caudale. La parte della testa di questo bastoncino, quella che sarà la testa, si ripiega al di sotto e all’indietro di 120° e la parte posteriore fa la stessa cosa. L’embrione assume una forma a C e si può propriamente parlare di embrione: l’embrione non è più asside ma ripiegato.
Durante le prime 9 settimane di gravidanza, infatti, si verificano una serie di trasformazioni significative nell’embrione e nel corpo materno.
- Settimane 1-2: Durante questo periodo, la blastocisti si impianta nell’endometrio. Le cellule germinali primordiali iniziano a formare i precursori degli organi.
- Settimane 3-4: L’embrione inizia a sviluppare il tubo neurale, che darà origine al sistema nervoso centrale. Si formano anche le prime cellule cardiache e il cuore inizia a battere. Iniziano a formarsi le prime strutture che diventeranno gli occhi e le orecchie.
- Settimane 5-6: L’embrione mostra i primi abbozzi di arti superiori e inferiori e si sviluppano ulteriormente il cuore e il sistema circolatorio. Inizia a formarsi quello che sarà l’apparato digerente.
- Settimane 7-8: L’embrione sta diventando un feto. Si formano le dita delle mani e dei piedi, mentre i reni iniziano a funzionare e l’embrione inizia a urinare nell’ambiente amniotico circostante.
- Settimane 9-11: In questa fase iniziamo a parlare di sviluppo fetale, è il momento in cui gli organi interni continuano a maturare.
In questo arco di tempo, alcuni nutrienti giocano un ruolo essenziale nel favorire il corretto sviluppo embrio-fetale, nell’attività della placenta e nella regolazione del flusso sanguigno. Uno studio della SUNY Downstate University di New York, pubblicato sullo speciale della rivista Nutrients, quest’anno, sottolinea l’importanza dei folati e delle vitamine del gruppo B, nello sviluppo neurologico nel periodo fetale fino all’età adulta di un individuo.
A 11 settimane di gravidanza parliamo di feto.
- Aspetto: Il viso inizia a svilupparsi ulteriormente, con l’apertura degli occhi. Le orecchie si spostano verso la loro posizione definitiva e le narici cominciano a distinguersi. Le dita delle mani e dei piedi hanno le unghie in formazione.
- Sistema nervoso: Continua a svilupparsi rapidamente. Il cervello produce sempre più neuroni e inizia a formarsi il midollo spinale. Il feto può fare movimenti involontari, sebbene la madre non li percepisca ancora.
- Organi interni: Continuano a maturare, infatti Il cuore del feto è completamente formato e batte regolarmente. Il fegato inizia a produrre bile e i reni cominciano a filtrare l’urina. L’apparato digerente è in fase di sviluppo.
- Sistema circolatorio del feto: Si sta sviluppando ancora meglio. Il cuore pompa il sangue attraverso i vasi sanguigni e il sangue inizia a trasportare ossigeno e nutrienti ai tessuti del feto.
- Dimensioni: Rispetto alle prime settimane di gravidanza, il feto a 11 settimane ha una struttura fisica più definita. Le parti del corpo stanno assumendo una loro forma, il sistema nervoso continua a evolversi, aprendo la strada ai futuri sviluppi cognitivi, e inizia a coordinare i movimenti. Questo rappresenta un momento particolare non solo a livello biologico ma proprio emozionale per la coppia, soprattutto quando attraverso gli esami strumentali è possibile ascoltare il battito cardiaco.
La Coltura Embrionaria in Vitro: Progressi nella Procreazione Medicalmente Assistita
Nell’ambito della Medicina della Riproduzione e della Procreazione Medicalmente Assistita (PMA), “embrione” e “blastocisti” sono senza dubbio tra i termini più utilizzati. Dagli albori della Procreazione Medicalmente Assistita, la tecnologia su cui ci si appoggia ha fatto passi da gigante. Le prestazioni vengono erogate in Regime Privato o convenzionato con Fondi Sanitari.
Per effettuare una coltura degli embrioni allo stadio di blastocisti, nell’ambito di un trattamento di PMA, è possibile ottenere in vitro lo sviluppo dell’embrione umano per un periodo massimo di sette giorni. Questo sviluppo in laboratorio consente di monitorare attentamente i fenomeni che portano a questa trasformazione, che sono molti, complessi ed estremamente delicati.

Durante un ciclo di Fecondazione in vitro (FIV), il trasferimento embrionale viene tipicamente eseguito proprio allo stadio di blastocisti. Il processo di differenziazione cellulare è cruciale per lo sviluppo dell’embrione, poiché determina quali cellule formeranno gli organi e i tessuti specifici. In un ciclo di FIV, il medico suggerirà il giorno in cui il trasferimento degli embrioni offre le maggiori possibilità di ottenere una gravidanza.
Le tecniche di coltura degli embrioni allo stadio di blastocisti si sono affinate nel tempo: tecnologia, elettronica e scienze dell’informazione hanno trasformato nei decenni i laboratori di PMA. In principio, i laboratori possedevano soltanto le attrezzature basilari, come incubatori, microscopi, centrifughe, bilance analitiche e poco altro. Ai giorni d’oggi gli stessi laboratori si sono trasformati. Incubatori e microscopi oggi sono strumenti evoluti che consentono di osservare e registrare in modalità time-lapse la fecondazione e lo sviluppo dell’embrione.
Progressi enormi sono stati ottenuti anche nella formulazione dei terreni di coltura embrionale. I primi terreni erano miscele colturali aspecifiche, le formulazioni erano spesso preparate nel laboratorio IVF, secondo criteri di qualità e riproducibilità non sempre ottimali. Esistono due principali approcci nella coltura: Sequenziale, che prevede la coltivazione degli embrioni in una serie di terreni usati in sequenza, a partire dalla fecondazione e per i successivi 5-6 giorni fino al trasferimento in utero; e Uso singolo, dove gli embrioni vengono coltivati in un singolo terreno.
Anche la tecnologia delle apparecchiature richieste per la coltura embrionale è progredita significativamente. I vecchi incubatori erano ingombranti contenitori nei quali il mantenimento e il monitoraggio delle condizioni di coltura ritenute necessarie per l’embrione (37°C e 20% CO2) era un’operazione ardua. Attualmente invece, gli incubatori sono alquanto compatti e dotati di accessori in grado di misurare in maniera continua e precisa temperatura e parametri micro-atmosferici, senza perturbare l’embrione durante la coltura. In particolare, studi sistematici hanno appurato che, soprattutto per la coltura allo stadio di blastocisti, una tensione parziale di CO2 al 5%, più vicina alle condizioni di relativa ipossia in vivo presenti nella tuba e nell’utero, sia nettamente da preferire a quella (20%) utilizzata in precedenza per decenni. Questa singola e semplice modifica assicura un incontestabile beneficio in termini di tassi di successo PMA (misurato come gravidanze evolutive per prelievo ovocitario). Per esempio, già agli inizi degli anni novanta del Novecento era possibile coltivare gli embrioni allo stadio di blastocisti con percentuali di sviluppo del 40% (percentuale esprimente il rapporto tra blastocisti ottenute rispetto al numero di ovociti fecondati). Tuttavia, in quei casi soltanto il 6-7% delle blastocisti ottenute e trasferite in utero erano in grado di impiantarsi.
tags: #blastocisti #annessi #embrionali