L'ibridismo rappresenta un insieme di fatti, di metodi e di teorie riguardanti gl'incroci tra forme differenti di animali e di piante. Il valore attribuito all'ibridismo come fenomeno biologico ha subito cambiamenti con lo sviluppo della scienza ed è in stretta relazione col significato attribuito in varî tempi all'espressione biologica "specie". Nella biologia moderna, dopo le ricerche del Mendel sulla trasmissione dei caratteri ereditarî, si chiamano ibridi (ibridi mendeliani) anche i prodotti dell'incrocio d'individui della stessa specie e razza, che differiscono per un sol carattere (monoibridi) o per due (diibridi) o tre (triibridi), ecc. In questo senso, tutte le fecondazioni che avvengono in natura fra gameti provenienti da individui diversi sono ibridazioni.
Un animale ibrido è un organismo nato dall’incrocio tra due specie differenti. Quando due specie sono evolutivamente vicine e non hanno ancora sviluppato barriere riproduttive (come differenze negli organi sessuali, incompatibilità genetica o rituali di accoppiamento diversificati), l’incrocio è ancora possibile e può dare origine a una progenie ibrida, che combina tratti fisici e comportamentali delle due specie parentali.
Il concetto di specie e la prospettiva storica
Giorgio Cuvier, il quale riteneva che le specie organiche siano stabili, ammetteva peraltro che il criterio dell'aspetto esteriore non è sufficiente a distinguere le vere specie originarie dalle varietà sorte per azioni esterne o per modificazioni di caratteri secondarî, non sostanziali, del plasma germinale. L'"incrocio" sarebbe il metodo che permette di compiere la distinzione: secondo il Cuvier due specie reali e diverse, incrociate fra loro, non danno prodotto, o lo dànno sterile, mentre due varietà della medesima specie, anche se molto differenti nell'aspetto morfologico, dànno prodotti fecondi.
Sebbene la maggioranza dei biologi, nella seconda metà del secolo passato, abbia seguito anche in questo la teoria darwiniana, gli zootecnici sono rimasti fedeli alla concezione del Cuvier al punto di distinguere due categorie d'incroci: gl'ibridi prodotti da due differenti specie e che sono sempre sterili, i meticci prodotti da due diverse razze o varietà, sempre fecondi. Esempio tipico del primo caso è il mulo, che nasce dall'incrocio dell'asino con la cavalla, del secondo il prodotto dell'incrocio fra due razze diverse di cani, di conigli o di polli. Tale distinzione degli zootecnici, se può avere un valore pratico, limitatamente alle pochissime specie di animali domestici ed eventualmente anche a quelle delle piante coltivate, non è di alcun aiuto per il sistematico.
Meccanismi di amissia e sterilità
L'osservazione che in molti casi l'unione è sterile e che in altri il prodotto di unione fertile ora è sempre sterile, ora è qualche volta fecondo, mentre talaltra è sempre fecondo, ha indotto i biologi di questo secolo a cercare le cause di queste differenze o per lo meno a stabilire quale sia il rapporto tra la fisiologia e la morfologia delle cellule germinali degl'ibridi. La fecondità completa coincide con l'affinità gametica, ossia con la capacità delle cellule germinali a produrre germi che sviluppano normalmente tutte le parti di un organismo, comprese le ghiandole germinali stesse ed i loro prodotti; la sterilità coincide con l'amissia, ossia con l'attributo opposto all'affinità gametica.
La coesistenza dell'affinità gametica con l'amissia ha luogo quando questa è dovuta a cause secondarie, somatiche o di ambiente, che non riguardano la capacità di formazione del germe da parte di due cellule germinali prodotte da forme differenti, anche se esse sono soltanto razze della medesima specie. Esiste un'amissia psichica, dovuta a una ripugnanza invincibile che una specie ha verso un'altra: tale è il caso dei bovini domestici e dei bufali, che non s'incrociano. Vi è un'amissia etica, derivante da costumi diversi: esistono alcune specie di pesci molto affini tra loro che possono, con accorgimenti artificiali, mescolarsi e dare origine a una discendenza feconda, ma normalmente i loro prodotti germinali non possono coniugarsi, perché emessi in diversa stagione o in ambiente diverso, come la superficie di un lago o il suo fondo.
Vi è finalmente un'amissia meccanica, dovuta all'impossibilità di penetrazione di spermî in uova provvedute di membrana più resistente di quella che appartiene alla specie donde derivano i primi o rivestite di guscio munito di apertura (micropilo) di diametro inferiore alle dimensioni dello spermio stesso. In questi ultimi casi ciò che non è possibile in un determinato senso è invece possibile nell'altro: il fatto è stato sperimentato in alcuni ricci di mare; per esempio gli spermî di Strongylocentrotus lividus fecondano facilmente le uova di Echinus microtuberculatus, ma non è possibile fecondare uova di Strongylocentrotus lividus con spermî di Echinus microtuberculatus per la ragione sopra indicata. Analogamente nei vegetali: Brassica oleracea non è fecondabile col polline di Brassica napus, mentre riesce l'incrocio inverso.
La barriera genetica e la speciazione
La nascita di una nuova specie è un processo che richiede milioni di anni e che, nella maggior parte dei casi, comincia con la separazione tra due gruppi di animali appartenenti a una stessa specie originaria. Facciamo un esempio concreto. I leoni e le tigri sono “specie sorelle”, che derivano cioè da uno stesso antenato comune, probabilmente vissuto in Asia in un’era geologica chiamata Miocene. Nel corso dei milioni di anni, complici alcuni cambiamenti climatici e ambientali, alcuni gruppi si separarono e migrarono in luoghi molto diversi. Un gruppo si stabilì in Africa, dove le ampie savane e praterie favorirono predatori specializzati nella caccia di gruppo e nella resistenza alla corsa. Da qui nacquero i leoni. Un altro gruppo si fermò in Asia, dove le foreste e le giungle selezionarono gli individui con corpo possente e mantello striato, ideale per mimetizzarsi tra la vegetazione. Da qui nacquero le tigri.
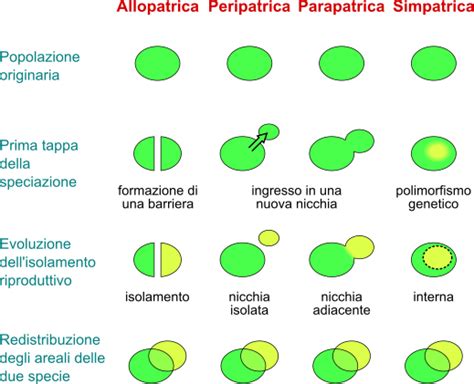
Nel corso delle ere geologiche, la formazione di barriere geografiche (catene montuose, innalzamento del livello dei mari, deserti) e l’adattamento a pressioni ambientali specifiche resero impossibile per i due gruppi incontrarsi nuovamente. Così, ciascun gruppo intraprese una storia evolutiva indipendente, fino a diventare due linee geneticamente distinte, ognuna delle quali, presa singolarmente, era costituita da un insieme di individui simili, capaci di incrociarsi liberamente all’interno del proprio gruppo e generare prole fertile. Questo processo, chiamato speciazione, è ciò che dà origine a quelle che oggi chiamiamo specie.
Quando due specie si sono separate da molto tempo, come per esempio cani e volpi, possono accumulare differenze genetiche tali da sviluppare delle vere e proprie barriere riproduttive (differenze negli organi sessuali, nei tempi e nei rituali di accoppiamento) che rendono impossibile la riproduzione. Tuttavia, se la speciazione è avvenuta in epoche più recenti, come nel caso della tigre e del leone, le due specie sono ancora abbastanza simili da potersi riprodurre e generare una prole. Questi animali ibridi sono spesso sterili, in quanto le differenze genetiche tra le specie parentali, prima fra tutte il diverso numero di cromosomi, non consentono la formazione di spermatozoi e cellule uovo vitali.
Classificazione biologica degli ibridi
L'amissia primaria, reale, gametica, è data dall'incapacità di due cellule germinali di sesso differente a coniugarsi e a produrre un germe. Sperimentalmente è stata osservata l'esistenza di germi ibridi fra specie molto differenti, i quali periscono poco dopo l'inizio della segmentazione dell'uovo. In altre lo sviluppo embrionale si arresta a stadî diversi; alcune dànno origine a larve che muoiono appena schiuse o prima di compiere la metamorfosi o in altri periodi dello sviluppo. Fin qui si potrebbe ancora parlare di amissia assoluta, per la mancata vitalità del prodotto.
Successivamente esiste una categoria di ibridi vitali e somaticamente vigorosi, i quali appartengono sempre al medesimo sesso e sono sempre infecondi, anche con le specie progenitrici. Citiamo, p. es. gl'ibridi del pavone e del gallo con la gallina di Faraone, che sono sempre maschi e completamente sterili. L'affinità gametica di due specie diverse s'inizia in forma saltuaria e accidentale, quando quelle generano ibridi normalmente sterili fra i quali tuttavia si osservano molto raramente individui fecondi in uno solo dei due sessi, individui capaci pertanto di procreare solo quando abbiano la possibilità di reincrociarsi con una delle specie pure dalle quali sono derivati.
Sotto questo aspetto il Poll ha distinto gl'ibridi in toconoti (da τόκος "parto" e νόϑος "bastardo") e steironoti (da στεῖρα "sterile" e νόϑος "bastardo"): i primi offrono la completa maturazione delle cellule germinali, nel senso che essi producono, rispettivamente nell'uno e nell'altro sesso, spermatozoi e uova mature. Tuttavia questo fenomeno non è indice assoluto e sicuro di fecondità, giacché esistono, in tale categoria d'ibridi, toconoti fecondi e toconoti sterili; la differenza è talvolta specifica e talaltra individuale, ma è sufficiente a dimostrare che non basta la completa maturità delle cellule germinali perché un ibrido riesca fecondo con altro individuo della stessa generazione o con altro appartenente ad una delle specie progenitrici.
Che cosa sono i cromosomi?
Poiché a partire dello stato di oogonio e spermatogonio le cellule germinali subiscono tre divisioni indirette (mitotiche) per giungere allo stadio di spermatozoo e di uovo maturo, l'autore citato chiama gl'ibridi toconoti col nome di trimitotici, i quali sono normali nella loro gametogenesi. Perché un ibrido raggiunga lo stato trimitotico bisogna che le due specie progenitrici abbiano apparati cromosomici e corredi fattoriali sufficientemente simili.
Negl'ibridi steironoti si distinguono diversi gradi:
- Apomitotici: mancano delle ultime tre mitosi di maturazione, così che nelle loro gonadi non si trovano che oogonî e spermatogonî.
- Monomitotici: ha luogo soltanto la prima mitosi che dà origine a spermatociti ed oociti di primo ordine.
Esempi notevoli e dinamiche di popolazione
I più comuni ibridi trimitotici ottenuti sperimentalmente sono quelli derivanti da incroci fra parecchie specie di fagiani, come il Dorato (Chrysolophus pictus) e quello di Lady Amherst (C. amherstiae), di Anatre, come Daphila acuta ed Anas boscas (Codone e Germano reale), di Fringillidi, come lucarino (Chrysomitris spinus) e canarino (Serinus canarius) fra gli Uccelli. Tra i Mammiferi invece gl'incroci del cane col lupo e con lo sciacallo, quelli di alcune specie di pecore fra loro, di alcuni Bovidi, ecc.
Tra gl'ibridi dimitotici vanno ascritti il comune incrocio fra l'Anatra comune (Anas boscas) e l'Anatra muta (Cairina moschata), gl'ibridi tra varî generi di anatre e di fagiani, il mulo e il bardotto, gli zebroidi, i maschi di parecchi incroci fra Bovidi come toro e bisonte, toro e jak. In conformità a quanto è stato osservato sperimentalmente, la maturazione delle cellule germinali non coincide nei due sessi di uno stesso ibrido: la gonomonarrenia e la gonomonoteliidia, sopra ricordate, corrispondono alla maturazione delle cellule germinali in un solo sesso.
Haldane, raccogliendo numerosi dati statistici, ha formulato il seguente principio: quando in una specie di ibrido uno dei due sessi manca o è raro o è sterile, questo è sempre il sesso eterozigotico, mentre il sesso presente o comune o fecondo è quello omozigotico. Abbiamo visto infatti che negli ibridi di Uccelli sono spesso fecondi i maschi, il cui sesso è omozigotico, mentre nei Mammiferi, nei quali è omozigotico il sesso femminile, sono spesso feconde le femmine, ed i maschi ibridi corrispondenti sono sterili. Così accade nei rari casi di fecondità delle mule e nella normale fecondità delle femmine ibride fra toro e bisonte e viceversa, fra toro e jak e viceversa.
L'ibridazione come forza evolutiva e alterazione antropica
L'uomo, fin da antico, ha cercato di produrre ibridi fra gli animali domestici e fra le piante coltivate, per interesse economico o per semplice diletto. Però molti ibridi nascono anche in natura, e poiché il loro aspetto è molto differente da quello delle specie progenitrici, i sistematici li hanno registrati e descritti. Gl'ibridi più comuni di animali domestici sono, oltre al mulo che nasce dalla cavalla e dell'asino, il bardotto che nasce dall'asina e del cavallo, lo chabin bastardo di capra e pecora, il leporide bastardo del coniglio con la lepre. Tra gli uccelli si conoscono ibridi dell'oca con l'anatra, del pavone con la gallina di Faraone, di questa col gallo, del gallo con la fagiana, del colombo con la tortora.
Si conoscono ibridi abbastanza numerosi tra animali domestici e specie selvatiche: il toro si incrocia col bisonte americano ed europeo, col yak, col banteng; la capra con lo stambecco; la pecora col muflone; il cavallo con la zebra; il maiale col cinghiale; il gatto domestico col selvatico; il cane con lo sciacallo, col lupo e con la volpe. La letteratura zoologica è ricca di citazioni e descrizioni di ibridi ottenuti tra varie specie selvatiche nei parchi, nei giardini, nelle uccelliere, negli acquarî e anche in natura. Numerose specie di farfalle e di coleotteri, di molluschi, di crostacei, di echinodermi sono state ibridate artificialmente e molti ibridi sono stati osservati direttamente.

Per ibridazione interspecifica si intende il processo per cui due animali di specie diverse si riproducono dando vita a una progenie con caratteristiche genetiche (e spesso, morfologiche) intermedie rispetto alle specie parentali. Tale progenie può essere sterile o fertile. In quest'ultimo caso, spesso, gli ibridi sono capaci di reincrociarsi con una o entrambe le specie parentali ("backcrossing"), costituendo un vettore per il passaggio di geni tra una specie e l'altra (introgressione). L'"ibridazione introgressiva" così risultante da un lato è una potente forza evolutiva, introducendo nuovi caratteri in popolazioni e specie, che possono o meno venire selezionati dall'ambiente; dall'altro, in alcuni casi, i nuovi caratteri introdotti in una specie possono risultare svantaggiosi e portare ad una riduzione della fitness e un aumento del rischio di estinzione.
Un caso particolarmente "problematico" riguarda l'ibridazione tra conspecifici domestici e selvatici. Poiché le pressioni selettive che agiscono sulle popolazioni in cattività sono molto diverse da quelle che agiscono sulle popolazioni selvatiche (rilassamento della pressione selettiva naturale, forte selezione artificiale), molto spesso i caratteri degli animali domestici, che possono passare alle popolazioni selvatiche tramite ibridazione introgressiva, risultano fortemente svantaggiosi. Un caso di ibridazione in natura può essere quello dell'incrocio tra l'orso polare e il grizzly, l'orso grolare. Sebbene, infatti, le due specie vivano in habitat diversi, lo scioglimento dei ghiacci artici sta spingendo gli orsi polari sempre più a sud, dove entrano in contatto con i grizzly. Da questi incontri possono nascere questi individui particolari, gli orsi grolari, con zampe marroni e mantello bianco.
Aspetti morfologici e complessità ereditaria
L'aspetto corporeo degl'ibridi è generalmente intermedio a quello dei genitori, tanto se si considerano le caratteristiche morfologiche, quanto se si studia il loro comportamento fisiologico. Poiché negl'incroci tra le razze e le varietà di una medesima specie di animali domestici o di piante coltivate i prodotti seguono le leggi dell'eredità alternata, bisogna premettere che queste ultime sono generalmente differenti per caratteri dovuti a un grande numero di fattori ereditarî. Pertanto, ove non accada che i fattori presenti in ciascuno dei genitori siano eterozigotici, nel qual caso si può attendere una prima generazione composta d'ibridi diseguali, si avrà una massa di prodotti piuttosto uniforme.
La seconda generazione prodotta da questi ibridi eterozigotici comprenderà necessariamente un numero immenso di combinazioni e di fenotipi. La segregazione, indubbiamente manifesta per la ricomparsa di caratteri eguali a quelli degli antenati, non può estrinsecarsi nella sua pienezza che nel caso in cui l'intera generazione comprenda un grandissimo numero d'individui, tanto maggiore quanto più numerose sono le differenze fattoriali che si sono trovate in presenza. Per dare un esempio della scarsa probabilità, quasi impossibilità reale, della ricomparsa dei modelli integrali degli avi, basterà ricordare l'esempio degl'ibridi tra fagiano dorato e fagiano di Lady Amherst. Si tratta di due buone specie, tanto diverse di colore almeno al sesso maschile, che qualsiasi occhio profano le distingue nettamente al primo sguardo: in schiavitù sono state incrociate e hanno dimostrato la più completa affinità gametica, tanto che i loro ibridi si sono riprodotti su larghissima scala.
Orbene, i maschi di queste due specie non presentano meno di 30 differenze fattoriali, il che significa che la seconda generazione deve offrire qualche miliardo di combinazioni, delle quali una sola corrisponderà alla formula esatta dell'uno o dell'altro antenato. Poiché una femmina di questi fagiani produce al massimo una ventina di uova all'anno, è praticamente impossibile riottenere i progenitori allo stato puro. Le probabilità crescono quando si tratti di colture vegetali che consentano di ottenere un numero enorme di esemplari. Così è accaduto al Baur ed al Lotsy nelle loro colture d'ibridi di bocca di leone. Bisogna inoltre tener conto dell'ineguale frequenza delle varie combinazioni, fra le quali predominano quelle maggiormente eterozigotiche e perciò più somiglianti alla prima generazione, e che dànno più facilmente l'impressione di forme intermedie.
Casi studio: Ligri, Tigoni e la gestione degli ibridi
Un esemplare di ligre, l’animale ibrido nato dall’incrocio tra leone e tigre, mostra come le dinamiche di cattività possano alterare i ritmi naturali. Quando un leone maschio si accoppia con una tigre femmina nasce il ligre: un felino con un mantello dorato simile a quello del leone, attraversato da striature che ricordano una tigre. Questi esemplari possono raggiungere dimensioni colossali. Nelle leonesse, infatti, alcuni geni limitano la crescita dei cuccioli, ma questa regolazione viene meno nell’incrocio con una tigre, consentendo ai ligri di raggiungere lunghezze superiori a 3,3 metri e un peso medio di oltre 400 kg.
Questi animali ibridi mostrano tratti comportamentali misti, che ne complicano l’interazione con le specie parentali. A questo si aggiungono frequenti problemi di salute legati alle loro dimensioni eccezionali. Per tali ragioni, l’allevamento e la riproduzione di ligri e tigoni sono oggi oggetto di forte contestazione da parte delle organizzazioni per la tutela degli animali. Similmente, il mulo, ibrido che deriva dall’incrocio tra un asino maschio e una cavalla, combina la corporatura robusta di un cavallo con la resistenza e la tenacia dell’asino, qualità che lo rendono capace di affrontare condizioni proibitive per molti altri animali da tiro. Per questo motivo, è utilizzato come animale da lavoro da almeno 3000 anni, venendo spesso impiegato per trasportare carichi pesanti, superiori a quelli che un cavallo di pari dimensioni potrebbe sostenere.
Quando invece il padre è un cavallo e la madre un’asina nasce il bardotto, un ibrido generalmente più piccolo del mulo, ma con caratteristiche simili in termini di resistenza e temperamento. Gli equidi, grazie alla loro stretta parentela evolutiva, possono dare origine anche ad altri ibridi curiosi, come lo zebrallo, derivante dall’incrocio tra zebra e cavallo, o lo zebrasino, nato dall’unione tra un maschio di zebra e un’asina. Questi esempi dimostrano come la sistematica si scontri spesso con la fluidità dei confini biologici, dove l'intervento umano, la pressione evolutiva e le barriere di fertilità definiscono il confine tra ciò che è specie e ciò che è ibrido.