La gestazione, spesso considerata da molti un miracolo, è in realtà resa possibile, dalla fecondazione al parto, grazie a complessi processi chimico-fisici e biologici. Al centro di questi processi vi è la placenta, un organo vascolare temporaneo e uno degli annessi embrio-fetali vitali, insieme al sacco vitellino, all'amnios, al cordone ombelicale, all'allantoide e al corion. L'importanza a livello tassonomico, evolutivo e di conseguenza funzionale di questo organo, attraverso il quale avvengono gli scambi tra il corpo materno e l'embrione, è fondamentale. La placenta umana è un organo che permette lo scambio gassoso e nutrizionale fra il feto e la madre nel corso della gravidanza. Essa collega il sistema circolatorio del concepito e della donna gravida e funge da "barriera" di separazione tra l'ambiente amniotico del feto e l'ambiente della cavità uterina.
I Primi Passi dello Sviluppo Embrionale e l'Impianto Uterino
Il percorso che porta alla formazione della placenta inizia con la fecondazione. Gli spermatozoi, gameti aploidi maschili, migrano attraverso il canale cervicale, la cavità uterina e nelle tube di Falloppio. Nell'ovaio si sviluppano i follicoli e, durante l'ovulazione, il follicolo dominante rilascia un ovocita, gamete aploide femminile. L'ovocita entra nella tuba di Falloppio attraverso l'estremità fimbriata, viaggiando attraverso di essa per poi passare nella cavità uterina. L'ovocita diventa un ovulo fecondato, o zigote, quando uno spermatozoo penetra negli strati esterni dell'ovocita. La fecondazione si verifica in genere quando l'ovocita si trova nell'ampolla della tuba uterina. Normalmente, un singolo spermatozoo fertilizza un singolo ovocita, mentre la fecondazione di due ovociti da parte di due spermatozoi porta a una gravidanza gemellare dizigotica.
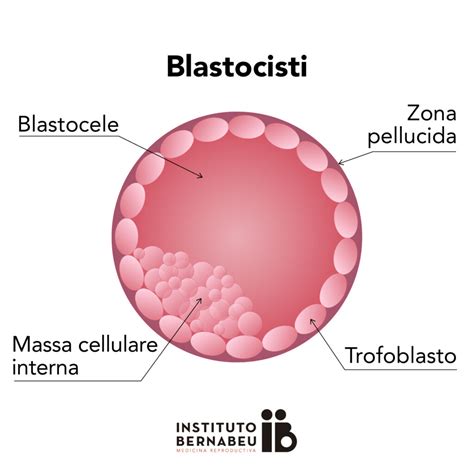
Lo zigote, diploide, si divide ripetutamente mentre migra verso il sito di impianto nell'endometrio, solitamente vicino al fondo o sulla parete posteriore dell'utero. Durante la sua migrazione nella tuba, l'uovo fecondato moltiplica i suoi blastomeri, processi definiti "segmentazioni", e si trasforma in morula, acquisendo diversi fenotipi. Successivamente, la morula si differenzia in un cumulo cellulare interno, l'embrioblasto, che costituisce il disco embrionale, e in uno strato cellulare periferico, il trofoblasto, deputato alla nutrizione dell'embrione. La morula si trasforma poi in blastocisti, caratterizzata da una cavità centrale ripiena di liquido detta blastocele. La blastocisti, attraverso la "seconda grande scelta differenziativa", darà origine a due tessuti extraembrionali: il sinciziotrofoblasto e il citotrofoblasto.
Circa sei giorni dopo la fecondazione, al momento dell'impianto, la blastocisti si annida nell'endometrio; il polo embrionale, che si svilupperà nell'embrione, è il primo punto dell'impianto. Con la disgregazione della zona pellucida, il sinciziotrofoblasto prende contatto con l'endometrio per apposizione, circa al 21° giorno del ciclo, con l'aiuto delle Matrix Metallo-proteinasi (MMP), dette anche matricine, che agiscono su integrine e matrice extra-cellulare (ECM). Le integrine sono glicoproteine transmembrana che collegano le proteine della matrice extracellulare ai microfilamenti intracitoplasmatici, costituendo un ponte che stabilizza il rapporto cellula-ECM e rende possibile la traduzione dei segnali dall'ECM al citoscheletro, e quindi un collegamento funzionale fra le cellule di un organo o tessuto. Nell'endometrio sono presenti 22 tipi di integrine e le principali sono vitronectina, fattore di Von Willebrand e osteopondina, mentre nell'embrione è presente la fibronectina. Per la stabilizzazione dell'annidamento è necessaria la scissione delle matrici extra-cellulari, e a tale scopo i trofoblasti secernono 22 tipi di enzimi specializzati, le Matrix Metallo-Proteinase (MMP), presenti sia sulle cellule endometriali che nell'embrione. Esse scindono le matrici extra-cellulari (ECM) della decidua permettendo la penetrazione del sinciziotrofoblasto e quindi l'impianto dell'embrione.
Il Ruolo Fondamentale dell'Endometrio e la Decidualizzazione
La porzione materna della placenta è denominata anche "pars basalis" ed è composta dalla tonaca mucosa (endometrio) dell'utero, quindi da un epitelio cilindrico semplice e da un connettivo sottostante. L'endometrio di una donna, durante il "periodo finestra" che consentirà l'annidamento della blastocisti, è soggetto a regolazione ormonale. Nello specifico, la formazione del corpo luteo in seguito all'ovulazione permette alla donna di possedere un livello ematico sempre più alto di progesterone. Da un punto di vista istologico il corpo luteo altro non è che un follicolo di Graaf trasformato. Grazie al progesterone, l'ovaio regola l'attività proliferativa delle cellule endometriali, inizializzando la fase proliferativa del ciclo mestruale. A livello molecolare, questo ormone si lega alle cellule stromali endometriali, inizializzando la reazione pre-deciduale: questi fibroblasti accumulano glicogeno e nutrienti; ne consegue che il connettivo endometriale diventa edematoso. Inoltre, il progesterone modifica la composizione del muco cervicale: da acido e denso diventa basico e meno viscoso, consentendo allo spermatozoo di risalire dall'utero verso la tuba uterina. I livelli di progesterone sono destinati a crollare in caso di mancata fecondazione, con la conseguenza che l'endometrio non sarà più una struttura adatta ad ospitare il futuro embrione.
Con l'impianto della blastocisti, l'endometrio uterino potrà diventare decidua. Prima di questa fase, che prende il nome di decidualizzazione, si verifica un altro periodo strettamente facente parte del ciclo uterino che prende il nome di predecidualizzazione, fase in cui lo strato funzionale dell'utero (endometrio) si prepara per divenire decidua. Infatti, in questa fase, l'endometrio assume e trattiene molto glicogeno e lipidi che saranno di fondamentale importanza per la nutrizione embrionale. In questa fase l'endometrio, sotto lo stimolo del progesterone, è in trasformazione deciduale: diventa iperplastico e le ghiandole aumentano di numero e di volume e secernono un liquido ricco di glicogeno e lipidi che forniranno nutrimento all'eventuale impianto della blastocisti. La decidualizzazione dell'endometrio è accelerata dall'impianto e rappresenta un meccanismo di difesa materna per impedire ulteriori processi di impianto. La decidua basale è situata immediatamente sotto il bottone embrionale.
Il Trofoblasto: Sinciziotrofoblasto e Citotrofoblasto
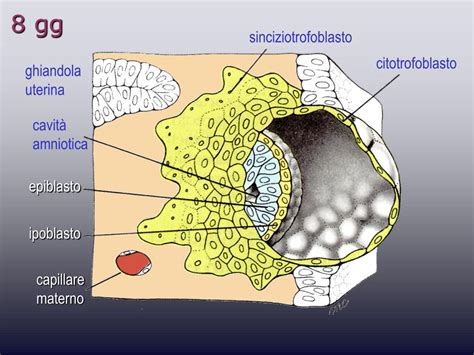
Entro uno o due giorni dall'impianto, uno strato di cellule, le cellule del trofoblasto, si sviluppa intorno alla blastocisti. Le cellule progenitrici dei villi del trofoblasto, ovvero le cellule staminali della placenta, si differenziano in due linee cellulari: il citotrofoblasto e il sinciziotrofoblasto.
Il sinciziotrofoblasto è un ammasso di cellule adibite all'erosione della mucosa uterina nella cosiddetta fase di impianto dell'embrione nei primissimi giorni di sviluppo dopo la fecondazione. Questa massa sinciziale (plasmoditrofoblasta) deriva dalla proliferazione di cellule, sottostanti ad essa, che costituiscono uno strato molto importante, il citotrofoblasto. Il tessuto sinciziale è una struttura "innovativa", non è presente nell'organismo se non nella placenta. È quella parte del trofoblasta che è venuta in diretto contatto col sangue materno e subisce una differenziazione. Il sinciziotrofoblasto continua ad accrescersi, diffondendosi nell'endometrio circostante, la "moquette" cellulare che riveste la superficie interna dell'utero, e creando soluzioni di continuità - dette lacune trofoblastiche - sempre più ampie. Riveste la superficie esterna del trofoblasto, copre i villi e sintetizza ormoni trofici, producendo gonadotropina corionica entro il decimo giorno e altri ormoni trofici poco dopo. Esso fornisce scambi arteriosi e venosi tra la circolazione embrionale e quella materna.
Il citotrofoblasto è lo strato profondo del trofoblasto che conserva la disposizione cellulare. Le cellule del citotrofoblasto penetrano, si approfondiscono nel sinciziotrofoblasto, a mo' di travate cellulari allungate. A sua volta, è sotteso da un tessuto definito mesoderma extraembrionale somatico. Il corion è costituito dal sincizio trofoblasto (parte esterna), dal citotrofoblasto (parte interna) e dal foglietto parietale del foglietto extraembrionale.
La Formazione e Struttura dei Villi Coriali
Mentre il trofoblasto si differenzia, emana gemme epiteliali che costituiscono i villi primitivi, conferendo all'uovo in questo periodo il suo aspetto villoso. Entro questi villi epiteliali, la lamina connettivale del corion manda i suoi prolungamenti che costituiranno lo stroma dei villi, e che riceveranno più tardi le ramificazioni dei vasi allantoidei portanti a ogni villo il sangue fetale. Circa cinque giorni dopo anche il mesoderma, tessuto embrionale primordiale, prolifera e si incunea nell’asse citotrofoblastico, producendo un’impalcatura vascolare, con capillari generati ex novo: il risultato finale sono i villi coriali, emanazioni longilinee che si proiettano esternamente, simili a tante dita, e che, come radici, gradualmente affondano nella compagine dell’utero, creando una fitta, intricata rete all’interno dell’endometrio.
Dal 10° al 14° giorno diventano visibili i villi primari, piccoli, non vascolarizzati, formati dal sinciziotrofoblasto superficiale e dal citotrofoblasto sottostante. Questi invadono il tessuto deciduale e le arteriole spirali, che nel frattempo hanno perso la tunica muscolare; l'invasione trofoblastica determina lo stravaso del sangue materno negli spazi intervillosi creando così le lacune ematiche. Successivamente i villi primari aumentano di volume e tra di essi si insinua un abbozzo di setto mesodermico proveniente dallo strato spongioso della decidua: si formano i villi secondari, che presentano al loro interno il mesoderma extraembrionale. Questi si trasformano in villi terziari, caratterizzati dalla presenza di setti deciduali ben definiti che dividono i villi in gruppi di cotiledoni. All'esterno i villi presentano un bordo a spazzola prominente (chorion frondosum), che gioca un ruolo chiave nei fenomeni di assorbimento dei gas di scambio e dei principi nutritivi. Ricerche eseguite con il microscopio elettronico hanno dimostrato che il bordo libero dei villi coriali presenta una serie di microvilli capaci di inglobare con i loro movimenti macromolecole proteiche o lipoproteiche.
La placenta si forma con la penetrazione dei villi coriali nell'endometrio. Queste strutture, inizialmente, si svilupperanno autonomamente e solo successivamente si andranno a unire per formare il primo abbozzo della placenta. La placenta primitiva si forma dalla fusione tra corion e decidua basale materna.
Anatomia Macroscopica della Placenta Matura
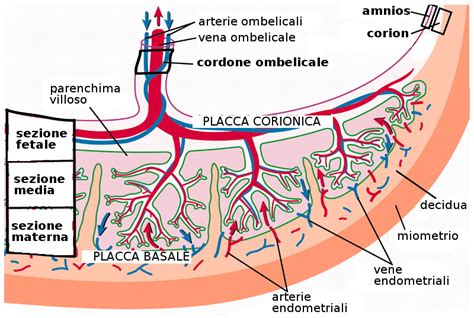
La placenta, organo discoidale, aderisce alla parete uterina per mezzo della caduca serotina. A mano a mano che l’organismo fetale si accresce, anche la placenta si estende, fino a costituire, al termine della gestazione, un disco di circa 20 cm di diametro. Essa è completamente formata entro le 18-20 settimane ma continua a crescere, fino a raggiungere il peso di circa 500 g al termine della gravidanza. Quando nel periodo del secondamento essa viene distaccata ed espulsa, si presenta in forma d'una focaccia, del diametro oscillante tra 16 e 20 cm, con uno spessore che decresce dal centro alla periferia, con un peso medio di circa 148 gr.
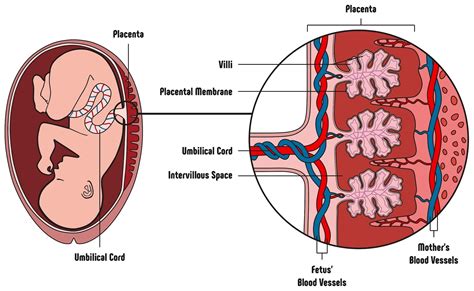
La placenta matura ha una faccia esterna o materna e una faccia interna o fetale. La faccia esterna, liberata dai coaguli sanguigni che vi aderiscono al momento della sua espulsione, ha un colore rosso scuro e presenta alcuni solchi che la dividono in un numero variabile di cotiledoni (da 15 a 20). Questi cotiledoni saranno divisi tra di loro esternamente da dei solchi intercotiledonali, internamente da dei setti intercotiledonali. Le due facce non si scolleranno tra di loro grazie a dei villi aderenti. L'altra tipologia di villi, presenti nelle lacune tra i villi aderenti, prendono il nome di "villi fluttuanti". La faccia interna è liscia e lucente, perché rivestita dalla membrana amnios, che lascia vedere per trasparenza le diramazioni dei vasi ombelicali sulla sottostante membrana corion. Il piatto amnio-coriale: le membrane fetali originano dal margine della placenta, sono formate da una sottile e lucente membrana amniotica e da un chorion più spesso ed opaco; sono separate dall’utero da una sottile decidua capsulare. Il sacco amniotico si riempie di liquido e si espande al crescere dell'embrione, riempiendo la cavità endometriale verso le 12 settimane circa dopo il concepimento; a questo punto, il sacco amniotico è la sola cavità rimanente nell'utero.
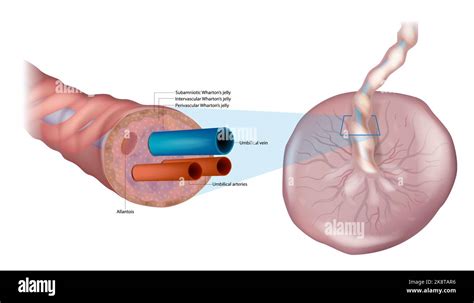
Il Cordone Ombelicale: Connessione Vitale
L'evoluzione del peduncolo d'attacco porta alla formazione del cordone ombelicale, l'unico collegamento fra l'embrione e la madre. A sviluppo definitivo le sue dimensioni raggiungono i 50 cm di lunghezza e risulta essere completamente rivestito dall'amnios. L'impianto del cordone ombelicale non è mai in posizione centrale rispetto alla placenta. L'unico inconveniente che il cordone ombelicale può dare è l'inserzione velamentosa, nel cui caso il cordone resta legato alla placenta solo con i vasi. Tale situazione non desta tuttavia problemi durante la gravidanza. Nella parte esterna il cordone ombelicale presenta dei rigonfiamenti denominati falsi nodi, causati dall'eccessiva lunghezza dei vasi rispetto al cordone. Visto in sezione appare "riempito" dalla gelatina di Wharton: un tessuto molle con poche fibre e molta sostanza cellulare, che costituisce l'impalcatura del cordone ombelicale. All'interno decorrono i vasi, in origine sono 4 (due arterie e due vene), in seguito le vene si fondono fra di loro. Nelle arterie circola sangue venoso che torna alla madre per l'eliminazione, nelle vene circola sangue arterioso ricco di nutrimento.
Le Molteplici Funzioni della Placenta
La placenta svolge un ruolo cruciale nella fisiologia materno-fetale. Le sue funzioni sono essenzialmente quelle di ossigenare, nutrire, difendere e disintossicare l'organismo durante lo sviluppo prenatale, oltre a possedere un'importante attività endocrina. La placenta è un organo temporaneo che, per via degli scambi e delle attività ormonali, svolge un ruolo sostitutivo di alcuni organi fetali non ancora attivi, come il polmone e i reni.
Scambio di Sostanze tra Madre e Feto
La placenta concorre agli scambi di sostanze fra madre e feto. Tra questi strati che separano nettamente il sangue materno dal sangue fetale si devono compiere tutti gli scambi necessari alla respirazione e alla nutrizione dell'uovo.Le più importanti funzioni fetali sono quelle della respirazione e della nutrizione. La placenta provvede alla funzione della respirazione fetale grazie agli scambi gassosi che si determinano tra sangue materno e sangue fetale attraverso i sepimenti che dividono le due circolazioni. L'ossigeno, che può così passare per diffusione osmotica, è quello solo che si trova disciolto nel plasma sanguigno, cioè una quantità esigua, assolutamente insufficiente all'ossigenazione dei tessuti fetali. La maggior parte dell'ossigeno del sangue materno è legato al globulo rosso nella forma di ossiemoglobina. Perché quest'ossigeno venga liberato occorre una disintegrazione della molecola dell'ossiemoglobina, occorre cioè una distruzione del globulo rosso. Questa distruzione è data dal potere citolitico, e più specialmente emolitico del trofoblasta, potere che si esercita incessantemente a livello della placenta per tutta la durata della gravidanza. Tale processo di emolisi è tanto intenso, che dovrebbe portare inevitabilmente all'anemia, se non fosse compensato da una più attiva rigenerazione dei globuli rossi negli organi emopoietici materni.Le sostanze di peso molecolare inferiore attraversano la barriera placentare con altre modalità, che richiedono in molti casi la partecipazione attiva del villo. Mentre per alcune sostanze (acqua, elettroliti, ossigeno, anidride carbonica, urea ecc.) gli scambi tramite la placenta avvengono in entrambe le direzioni in rapporto a fattori di ordine fisico-chimico (pressione idrostatica, pressione osmotica, pressione oncotica, diversa concentrazione esistente nei due versanti ecc.), per altre intervengono meccanismi biochimici di natura enzimatica, che regolano la direzione e la velocità degli scambi stessi.La placenta provvede anche alla nutrizione del feto. I materiali nutritizi a esso necessari non possono passare attraverso i setti che separano le due circolazioni se non allo stato di soluzione. Certo è che il feto non può ricavare i materiali di nutrizione a lui necessari che dalla placenta, per lo meno nei mesi più avanzati di gravidanza; solo nei primissimi giorni dall'incubazione il materiale nutritizio potrebbe essere fornito dalla vescicola ombelicale. Quanto al liquido amniotico, che il feto deglutisce continuamente, è troppo povero di materiali nutritizi per poter bastare alla sua nutrizione. Solo l'acqua, che è il componente principale del liquido amniotico, passa dallo stomaco del feto nella corrente sanguigna, dove concorre a diluire opportunamente il sangue e variarne la tensione osmotica, per regolare gli scambi osmotici attraverso alla placenta.Per quanto riguarda il materiale proteico, il transito è consentito solo a proteine a basso peso molecolare, in grado di attraversare la placenta. Tale aspetto è importante se si considera che buona parte degli ormoni sono di natura proteica ed a basso peso molecolare. Anche gli anticorpi, costituiti da proteine, migrano dalla madre al feto. Ma solo quelli aventi basso peso molecolare.
La PLACENTA PREVIA, una barriera impenetrabile - Spiegazione
Funzione Endocrina della Placenta
Tra i vari ruoli svolti dalla placenta, uno dei più affascinanti è la regolazione ormonale. La placenta è una ghiandola endocrina, in grado di produrre gonadotropine corioniche, ormoni con la funzione di mantenere attivo il corpo luteo (ghiandola endocrina che si forma nell'ovaio dopo l'ovulazione). Il corpo luteo produce progesterone implicato nel procedere della gravidanza. L'hCG, gonadotropina corionica umana, un ormone molto simile all'LH, è fabbricata dalle cellule del sinciziotrofoblasto e compare nel sangue materno subito dopo l'impianto: serve a preservare l'integrità del corpo luteo, presente a livello ovarico, da cui dipende la produzione del fondamentale progesterone. L'hCG può essere rintracciato nel sangue durante la prima settimana dopo la fecondazione, mentre i comuni test di gravidanza sulle urine, per uso casalingo, possono rilevare l'ormone un paio di settimane dopo. Se il suo livello di produzione dovesse calare ci sarebbe un'interruzione di gravidanza.
Evolvendosi, anche la placenta sarà in grado di produrre progesterone e, fra il terzo e il quarto mese di gravidanza, avrà raggiunto dimensioni tali da essere in grado di produrre autonomamente la giusta quantità di ormone. Nel contempo, la quantità prodotta dal corpo luteo tenderà a diminuire proporzionalmente. Se tale passaggio di ruolo tra placenta e corpo luteo non dovesse avvenire, il progesterone avrebbe un brusco calo di produzione con conseguente aborto spontaneo. Questa è la ragione per la quale il periodo più critico per la gravidanza è rappresentato proprio dal terzo-quarto mese.
La placenta, secondo le moderne vedute, va considerata anche come organo a secrezione interna, produttore di quegli ormoni che, passati nel circolo materno, vengono poi eliminati con le urine, dove la loro presenza è facilmente rivelata dalla reazione di Aschheim-Zondeck. La placenta produce numerosi ormoni e sostanze che abitualmente riconoscono altre sedi di origine in condizioni di non gravidanza. In tal modo, la placenta assume funzioni molto simili all'ipotalamo, ipofisi, fegato, surrene, ovaio e corpo luteo. Queste sostanze regolano il meccanismo immunitario dell'impianto, il metabolismo e la crescita fetale, lo scambio gassoso feto-materno e la contrattilità uterina. Tra questi vi sono estrogeni, ormone lattogeno placentare e Gn-RH, un neuropeptide placentare simile a quello ipotalamico. La sua secrezione e concentrazione plasmatica aumenta parallelamente a quella dell'HCG.
Fino a pochi decenni fa, la placenta era considerata un organo steroidogenetico completo, cioè capace di sintetizzare gli estrogeni e il progesterone a partire da precursori semplici. Oggi si è dovuto modificare tale concetto per quanto riguarda la steroidogenesi: la placenta è soprattutto un organo di ricambio e non di produzione. Essa accumula e soprattutto modifica steroidi preesistenti in circolo, prodotti dagli organi materni e fetali, grazie alla presenza di taluni enzimi (deidrogenasi, idrossilasi, solfatasi) molto attivi in essa presenti. Esistendo poi una certa diversa distribuzione di tali enzimi tra feto e placenta, le attività steroidogenetiche risultano per così dire integrate.
Funzione di Difesa e Barriera Placentare
Oltre alla funzione della respirazione e a quella della nutrizione, alla placenta spetta un'altra importantissima funzione fetale, quella della difesa del feto contro microrganismi patogeni che eventualmente circolassero nel sangue della donna malata. L'infezione del feto attraverso alla placenta è infatti da ritenersi fatto assolutamente eccezionale, che si verifica solo se il tessuto placentare abbia perduto per malattia questi suoi poteri di difesa. Per dire solo delle malattie infettive più comuni, è noto che il bacillo della tubercolosi non passa, si può dire, mai dalla madre al feto, né il plasmodio della malaria, né altri germi patogeni.Probabilmente la placenta non limita la sua funzione di difesa a impedire il passaggio di germi patogeni. Essa è in grado di trattenere e fissare anche talune sostanze che per la loro solubilità e diffusibilità si direbbe debbano passare facilmente attraverso il filtro placentare. Così in un gran numero di casi d'itterizia della gravida, i pigmenti biliari circolanti nel sangue materno non passano attraverso la placenta; tanto è vero che il feto non è itterico, mentre il tessuto della placenta si presenta intensamente colorato in verde. Questa capacità di fissare e trattenere i pigmenti biliari la si vuole attribuire alla presenza nella placenta di sali di calcio. È presumibile che una simile azione elettiva possa esercitarsi da parte della placenta anche riguardo ad altre sostanze, solubili o d'altro genere, circolanti nel sangue della gravida malata.Ad esempio, pertosse e varicella non sono in grado di attraversare la placenta. Tuttavia, durante il parto i virus e alcuni protisti (ad esempio alcune spirochete agenti della sifilide) passano dalla madre al feto. Anche buona parte dei farmaci compiono questa migrazione.
Circolazione Materno-Fetale e l'Integrazione Placentare
Quando l'uovo fecondato arriva nella cavità uterina per annidarsi nello spessore della mucosa, ha raggiunto lo stadio di gastrula, e rappresenta una formazione vescicolare, circondata tutto all'intorno da uno spesso strato epiteliale o ectodermico, detto trofoblasta. Al trofoblasta compete la singolare proprietà citolitica, la proprietà cioè di aggredire e disintegrare gli elementi con cui venga a contatto. Per questa sua proprietà, dovuta verosimilmente alla presenza di fermenti citolitici, l'uovo, che poggia sul rivestimento epiteliale della mucosa uterina, ne distrugge gli elementi, e si mette così in contatto diretto con gli elementi del sottostante stroma connettivale. Anche questi elementi vengono a loro volta aggrediti e distrutti dal trofoblasta, e così l'uovo si scava nello spessore della mucosa una cavità o nido, la camera incubatrice.Le pareti dei vasi della mucosa subiscono a loro volta l'azione aggressiva del trofoblasta, e allora nella camera incubatrice si versa, attorno all'uovo, sangue materno. Questo sangue non coagula, perché il trofoblasta con cui viene a contatto ha la stessa proprietà degli endoteli, quella cioè di mantenere fluido il sangue, che uscito in tal caso dalle piccole arterie erose, rientra per le vene del pari erose. Da questo momento l'uovo si trova immerso nel sangue che ne irrora tutta la periferia spingendosi fra i meandri del trofoblasta, e le propagini del trofoblasta vi rimarranno immerse per tutta la durata della gravidanza. Si ha dunque ragione di dire che l'uovo vive nel sangue materno.Al di fuori del villo sta il sangue materno; dentro il villo il sangue fetale; tra essi interposto il trofoblasta (plasmoditrofoblasta e citotrofoblasta), lo stroma connettivale del villo e le pareti dei vasi del villo. La placenta umana è di tipo emocoriale, vale a dire che il corion, impiantandosi nell'utero, corrode l'epitelio, il connettivo sino a raggiungere i vasi sanguigni materni.L'uovo si nutre per mezzo del deutoplasma con l'aiuto della secrezione derivante dalle ghiandole uterine che producono glicogeno. Durante l'annidamento il deutoplasma non è più sufficiente a fornire il nutrimento, spingendo il germe a nutrirsi dei tessuti materni che sono digeriti durante l'impianto (stadio isotrofico). Ad annidamento ultimato la nutrizione diviene di tipo emodrofico.La circolazione fetale è altamente specializzata. L'arteria ombelicale (contenuta nel cordone ombelicale), che arriva al fegato fetale. Il sistema cardiovascolare fetale presenta circuiti modificati rispetto a quello post-natale. Il 50% del sangue nel ventricolo destro va al polmone e 50% nel ventricolo destro. Solo il 10% raggiunge l'aorta discendente. Il sangue venoso ricco di ossigeno e nutrienti passa dalla placenta, dove cede i cataboliti e l'anidride carbonica al sangue materno.
La PLACENTA PREVIA, una barriera impenetrabile - Spiegazione
Classificazione e Varietà Morfologiche della Placenta
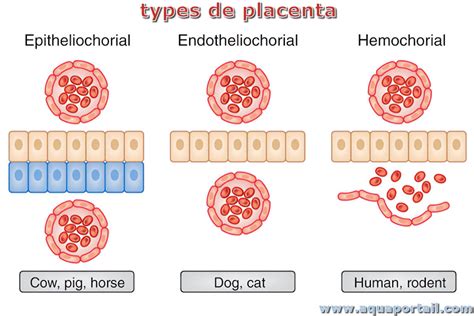
Le placente nei vertebrati vivipari presentano due tipi embriologicamente diversi: la placenta propriamente detta, propria dei Mammiferi Placentali e di taluni Rettili, costituita da una fusione tra il corion e l'allantoide e irrorata dai vasi allantoidei; la placenta vitellina, propria di taluni Squali e di taluni Rettili, formata in rapporto al sacco del tuorlo, cui nei Rettili si unisce il corion, e irrorata dai vasi vitellini. Presenta interesse il fatto che, in talune specie di Rettili, le due placente possono coesistere, onde il medesimo embrione ha placenta vitellina e placenta p.Il Bonnet distingue, in corrispondenza della placenta, due sorta di processi per la nutrizione embrionale: la nutrizione per embriotrofo, dove l'epitelio uterino elabora sostanze destinate all'embrione, che le assorbe per mezzo dell'epitelio coriale, e la nutrizione per emotrofo, dove le sostanze nutritizie passano dal plasma del sangue materno al plasma del sangue fetale per un semplice fenomeno di diffusione attraverso i tessuti interposti, ridotti a costituire una sottilissima membranella.La nutrizione per embriotrofo si verifica nelle zone di placenta in cui epitelio uterino ed epitelio coriale sono normalmente sviluppati (rapporto epiteliocoriale), o nelle quali, in punti limitati, sparisce l'epitelio uterino e il connettivo sottostante aderisce strettamente all'epitelio coriale (rapporto sindesmocoriale). Nelle placente dei Selaci e dei Rettili non si osserva una relazione emocoriale, ma le altre tre forme. Nei Mammiferi Placentali, dalle forme inferiori alle superiori, si vedono complicarsi gradatamente tali relazioni fino a giungere al rapporto emocoriale dei Primati.Il corion, nella zona in cui si forma la placenta, presenta lunghi villi riccamente irrorati, che si spingono entro cripte dell'epitelio uterino. Nei Mammiferi Placentali inferiori tutta la superficie del corion porta villi (placenta diffusa). Nei Ruminanti si trovano invece ciuffi di villi (cotiledoni) sparsi sulla superficie del corion, la quale, negli spazi tra essi, appare liscia (placenta cotiledonare). In tutti questi animali si osservano principalmente relazioni, tra tessuto materno e fetale, epiteliocoriali e sindesmocoriali; prevale cioè la nutrizione per emotrofo e vi è una scarsa aderenza tra tessuti materni e fetali, onde, dopo il parto, non vengono espulse decidue paragonabili alle decidue dei Mammiferi superiori e dell'uomo.Nei Carnivori la zona del corion, munita di villi, forma un anello intorno al sacco coriale (placenta zonaria). Nei Primati, Roditori, Insettivori, Chirotteri si osserva una placenta ristretta a una piccola zona del corion, la quale manda però villi molto lunghi che entrano profondamente nel tessuto uterino (placenta discoidale). In tutti questi animali si osserva il rapporto endoteliocoriale o perfino emocoriale. Prevale nutrizione per emotrofo, e vi è forte compenetrazione fra tessuti materni e fetali, onde dopo il parto vengono espulse decidue e il parto stesso è accompagnato da notevole emorragia materna.
Anomalie Morfologiche e Condizioni Patologiche della Placenta
La configurazione e la dimensione globale della placenta dipendono dalla precisa distribuzione e dallo stato di sviluppo e di funzionamento dei villi coriali. Le anormalità morfologiche della placenta sono da mettere in relazione al luogo definitivo di impianto dell’uovo fecondato in utero.La placenta può presentare diverse anomalie strutturali, tra cui:
- Placenta bi- tri- multi-partita (lobata): una divisione incompleta della placenta in due, tre o più lobi.
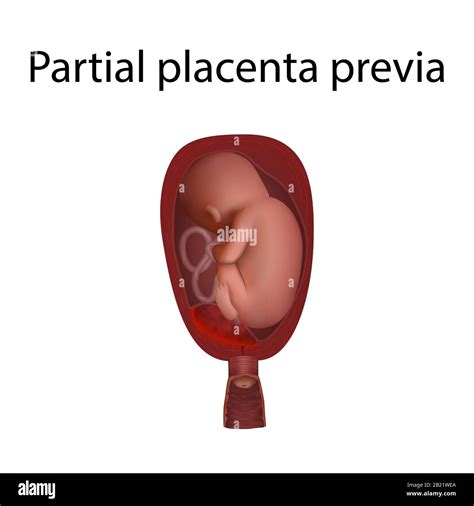
- Placenta succenturiata: una completa divisione della placenta in una massa centrale e uno o più piccoli lobi. L'impianto basso del lobo accessorio può dar luogo a placenta previa (vasa previa).
- Placenta membranacea: condizione in cui i villi coriali funzionanti includono il sacco amniotico nella sua interezza. È un evento raro correlato a un accrescimento fetale intrauterino ritardato e dal travaglio prematuro.
- Placenta circumvallata: la placenta presenta un piatto coriale ristretto dovuto alla reduplicazione delle membrane fetali lungo il margine periferico che può essere completamente circondato.
- Placenta circummarginata: il margine della placenta si proietta al di là dell'inserzione delle membrane fetali, così è possibile osservare un tessuto placentare che rimane non ricoperto. Questo tipo di alterazione strutturale si riscontra frequentemente nelle placente di gemelli.
La posizione dipende dal punto in cui si impianta l'embrione durante la "finestra d'impianto" (ottimale per un concepimento tra la 19esima e la 24esima giornata del ciclo uterino). Fisiologicamente la placenta potrà formarsi adesa alla parete posteriore o anteriore dell'utero, lateralmente destra o sinistra, o fundica (cioè alla porzione apicale dell'utero, il fondo). Se la placenta dovesse coprire parzialmente o completamente l'orifizio uterino interno si chiamerà placenta previa. Questa si forma dopo il corpo dell'utero, all'altezza del collo di questo, ed è presente nel 3% delle gravidanze singole. Questa può indurre mortalità materna, emorragia antepartum, intrapartum o postpartum, isterectomia e sepsi.
Normalmente la placenta si distacca dalla parete uterina attraverso lo strato fibrinoide di Nitabuch situato nello strato spongioso della decidua basale. Approssimativamente si perdono 250 cc di sangue durante il secondamento. Il parto con placenta di tipo emocoriale, tipica degli umani, delle scimmie antropomorfe e dei roditori, è particolarmente emorragico, a differenza di quello di buona parte delle altre specie animali.
Complicazioni e Patologie Placentari Specifiche
Le gravidanze gemellari hanno maggiori complicazioni perinatali specialmente nel caso di placente monocoriali monoamniotiche. Ci può essere tensione di uno o entrambi i cordoni ombelicali intorno alle varie parti del feto con ostruzione del flusso sanguigno. Occasionalmente si verifica l'amputazione di una estremità fetale. Nei gemelli con placente monocoriali si può riscontrare una fistola artero-venosa che mette in comunicazione i cotiledoni fetali, il sangue arterioso di un feto in tal caso rifornisce uno o più cotiledoni comuni, mentre questi sono drenati da vasi venosi verso l'altro gemello. Questo comporta che un gemello presenta policitemia e polidramnios mentre l'altro gemello presenta anemia e oligoidramnios.
L'eritroblastosi fetale può provocare un ingrandimento della placenta nei casi gravi di sensibilizzazione materna. L'idrope fetale invariabilmente presenta una placenta ingrandita che è di colorito variabile dal grigiastro al rossastro e relativamente compatta. L'esame microscopico rivela alcune divisioni irregolari nell'arborizzazione dei villi con aspetti a clava cui si associa la presenza di fibrosi e di cellule di Hofbauer (istiociti) e persistenza del citotrofoblasto di Langhans. I vasi sanguigni fetali possono mostrare eritrociti nucleati.
La placenta piccola può essere causata dalla prematurità. Un ridotto volume placentare equivale a scarsa irrorazione ed è predittivo di IUGR (Restrizione della Crescita Intrauterina) specialmente nel secondo trimestre.
La corionite o corionamnionite, una flogosi significativa del chorion e dell'amnios, si può riscontrare, insieme ad iperpiressia materna, nella rottura prolungata delle membrane fetali. Nella sifilide congenita, la placenta è di solito ingrandita, pastosa, di colorito giallastro o grigiastro con cotiledoni friabili. L'esame microscopico mostra i villi coriali a forma di clava con arborizzazione irregolare.
Gli infarti placentari hanno forma conica, sono compatti, di colore rosso, grigio o biancastro con contorni netti relativamente definiti. Essi sono causati dall'ischemia dei villi corionali, per trombosi dai vasi sanguigni fetali che penetrano nei cotiledoni fetali. Microscopicamente è presente una necrosi coagulativa dei villi con depositi intervillosi di fibrina e marcate lesioni emorragiche.
Nella gravidanza oltre il termine si realizza il quadro della cosiddetta senescenza placentare, che sta ad indicare il perduto optimum di funzionalità raggiunto con la maturazione. Ecograficamente possiamo stabilire un grading 0-3 corrispondente alle capacità funzionali placentari. Grado 3: presenza di echi francamente iperriflettenti ed irregolari da attribuire ad aree di calcificazione dei setti interlobari. Il 50% delle placente mostrano calcificazioni dalla 33a settimana. Lo spessore della placenta supera i 5 cm per edema e presenza di calcificazioni. È associato con IUGR, ipertensione arteriosa e fumo. Questa senescenza porta a una diminuzione della steroidogenesi (progesterone, estriolo e HPL): il progesterone ematico materno e il pregnandiolo urinario materno dimostreranno una sensibile diminuzione, come pure il dosaggio dell'estriolo, fornirà dei valori di molto inferiori alla norma in correlazione con la senescenza placentare dato che la conversione del DHEA (prodotto nel surrene fetale) in estriolo, avviene nella placenta.
tags: #placenta #epitelio #sincizio #trofoblasto