L'ecosistema botanico terrestre è straordinariamente ricco di forme di vita, ognuna con le proprie peculiarità evolutive e adattamenti unici. Tra queste, un ruolo di notevole interesse è rivestito dalle Pteridophyte, un gruppo di piante che, per la loro antichissima origine, rappresentano una testimonianza vivente delle prime fasi dell'evoluzione vegetale sulla terraferma. All'interno di questa vasta e diversificata divisione, il genere Equisetum si distingue per la sua morfologia inconfondibile e per un ciclo vitale che evoca tempi geologici remoti. La specie Equisetum arvense, comunemente conosciuta come Equiseto dei campi o Coda di cavallo, è una delle rappresentanti più diffuse e studiate di questo genere. Questa pianta perenne erbacea, che colonizza un'ampia varietà di habitat, presenta una dicotomia strutturale fondamentale tra fusti fertili e fusti sterili, ciascuno con ruoli e caratteristiche distintive. L'analisi del fusto fertile, in particolare, ci permette di addentrarci nelle strategie riproduttive di questa specie, rivelando una serie di adattamenti morfologici e fisiologici intrinsecamente legati alla disseminazione delle sue spore.
Classificazione, Nomenclatura e Patrimonio Genetico
L'Equisetum arvense appartiene alla grande divisione delle Pteridophyte, un gruppo di piante più primitive rispetto alle Angiosperme. Queste piante si caratterizzano per l'assenza di organi sessuali distinti nel senso tradizionale e si propagano e riproducono principalmente per mezzo di spore. Il genere Equisetum è ulteriormente suddiviso in due sezioni principali: Hippochaete e Euequisetum, riflettendo una certa diversità morfologica e ecologica al suo interno.
Il nome generico, Equisetum, deriva dalla lingua latina e si traduce letteralmente con “crine di cavallo”, un'evocazione diretta e pittoresca della tipica conformazione dei fusti sterili di molte specie, che ricordano per l'appunto la criniera di un equino. L'epiteto specifico, “arvense”, anch'esso di origine latina, fa esplicito riferimento al suo habitat tipico: i campi coltivati o incolti, i margini dei sentieri e le aree ruderali, indicando una predilezione per ambienti aperti e spesso antropizzati.
Nel corso della storia botanica, la specie Equisetum arvense ha conosciuto diverse nomenclature, riflettendo le continue revisioni e approfondimenti tassonomici. Tra le denominazioni sinonimiche che si sono succedute nel tempo, si annoverano Equisetum arcticum Rupr., Equisetum boreale Brongn., Equisetum campestre C.F., e Equisetum telmateia Ehrh. Questa molteplicità di nomi testimonia la complessità della sua identificazione in contesti diversi e l'evoluzione della comprensione scientifica delle sue relazioni filogenetiche. Un dato scientifico di particolare interesse, che sottolinea l'antica eredità e la robustezza genetica di questa specie, è il suo elevato numero di diploidi: 216, corrispondenti a 108 copie di cromosomi. Tale complessità genomica può essere interpretata come un indicatore della sua resilienza evolutiva e della sua capacità di adattarsi a una vasta gamma di condizioni ambientali nel corso di milioni di anni.
Il Ciclo Biologico Perenne e la Parte Ipogea: Geofita Rizomatosa
La forma biologica della specie è classificata come geofita rizomatosa (G rhiz), una categoria che descrive piante perenni erbacee le cui gemme, durante la stagione avversa, si trovano in una posizione sotterranea. Questo adattamento consente alla pianta di sopravvivere a condizioni climatiche sfavorevoli, quali gelo invernale o siccità estiva, ritirando completamente gli organi aerei e proteggendo i meristemi di crescita. Le gemme, da cui si dipartiranno i nuovi fusti epigei (aerei) nella stagione successiva, sono ospitate in organi sotterranei specifici, noti come rizomi. Un rizoma è un fusto ipogeo orizzontale, lungamente strisciante, che svolge un ruolo cruciale nella propagazione vegetativa e nell'accumulo di sostanze di riserva.
Da questo sistema rizomatoso, ogni anno, si dipartono nuove radici e fusti aerei, assicurando la continuità della specie nel tempo. La strategia di sopravvivenza dell'Equisetum arvense è ulteriormente perfezionata dalla presenza di ingrossamenti tuberiformi sui rizomi. Questi tuberi sono il risultato della trasformazione di brevi rami ipogei laterali e fungono da veri e propri magazzini di sostanze nutritive. Durante i periodi più avversi, quando le risorse esterne scarseggiano, la pianta attinge a queste riserve energetiche per mantenere la propria vitalità, permettendo al ciclo biologico di rimanere perenne e assicurando la ripresa vegetativa con l'arrivo di condizioni più favorevoli. Le radici, di tipo avventizio, sono secondarie e fascicolate, sviluppandosi direttamente dal rizoma, consolidando ulteriormente l'ancoraggio della pianta al substrato e la sua capacità di assorbire acqua e nutrienti dal terreno.
La parte ipogea del fusto, costituita da questo intricato sistema di rizomi orizzontali, ingrossamenti tuberiformi e varie ramificazioni, talvolta anche molto intricate, rappresenta la spina dorsale della pianta. È da questa rete sotterranea che si originano i germogli aerei eretti, i quali daranno vita ai caratteristici fusti epigei, sia sterili che fertili, che emergono dal suolo per svolgere le loro specifiche funzioni biologiche. Questa complessa organizzazione sotterranea non solo garantisce la sopravvivenza pluriennale della pianta, ma contribuisce anche alla sua notevole capacità di colonizzare e persistere in ambienti diversi, rendendola una specie robusta e tenace.
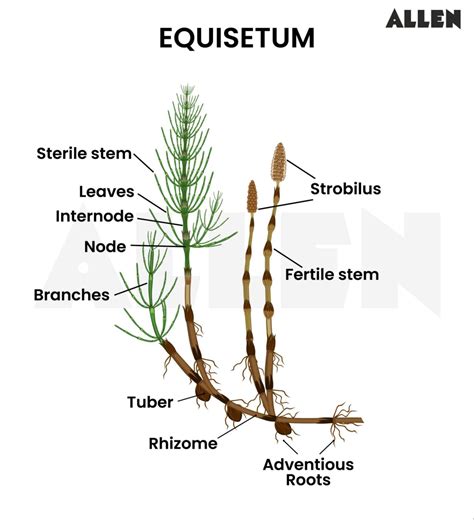
Morfologia dei Fusti: Il Contesto del Fusto Fertile
L'Equisetum arvense manifesta una notevole differenziazione morfologica tra i suoi fusti, che possono essere principalmente di due tipi: fertili e sterili. Questa distinzione è fondamentale per comprendere appieno la strategia riproduttiva e metabolica della pianta.
I Fusti Fertili: Pionieri della Riproduzione
I fusti fertili, l'oggetto centrale di questa trattazione, emergono tipicamente all'inizio della primavera, precedendo la comparsa dei fusti sterili. Essi sono strutturati principalmente per la riproduzione. Normalmente, questi fusti sono eterotrofi, il che significa che non svolgono la funzione fotosintetica in maniera significativa. La loro colorazione tende a essere più chiara, spesso brunastra o rossastra, a causa della scarsa presenza di clorofilla. Le loro dimensioni sono contenute, con un'altezza che varia tipicamente tra i 10 e i 20 cm. Questa limitata altezza permette loro di emergere rapidamente dal terreno, completare il ciclo riproduttivo e rilasciare le spore prima che la vegetazione circostante diventi troppo densa o che le risorse nutritive necessarie per la fotosintesi siano pienamente disponibili.
Nonostante la loro natura prevalentemente eterotrofa, in alcuni casi particolari si possono trovare individui con i fusti fertili provvisti di rametti verdi, seppur ridotti. In queste rare eccezioni, i fusti fertili possono acquisire una certa capacità fotosintetica, suggerendo una plasticità adattativa anche in questa struttura specializzata. Tuttavia, la loro funzione primaria rimane l'elevazione e il supporto dello strobilo, l'organo riproduttivo apicale che contiene le spore. La breve durata della loro vita, che termina una volta completata la disseminazione delle spore, sottolinea la loro specializzazione temporanea e riproduttiva.
I Fusti Sterili: Architetti della Fotosintesi
Al contrario, i fusti sterili emergono in un secondo momento, dopo che i fusti fertili hanno completato la loro missione. Essi sono i principali attori della fotosintesi per la pianta. Questi fusti sono ruvidi al tatto e di colore verde intenso, essendo ricchi di clorofilla, il pigmento essenziale per la conversione dell'energia solare in zuccheri. Le loro dimensioni sono generalmente maggiori, raggiungendo altezze tra i 20 e i 50 cm.
La colorazione dei fusti sterili può variare sensibilmente a seconda dell'ambiente circostante, riflettendo la capacità della pianta di adattarsi alle condizioni luminose e idriche. Nella zona apicale, possono apparire più arrossati in ambienti aridi, una pigmentazione che potrebbe svolgere un ruolo protettivo contro l'eccessiva irradiazione solare o lo stress idrico. Se l'ambiente è invece umido o scarsamente illuminato, i fusti tendono ad acquisire un aspetto verdastro-gialliccio, fenomeno noto come clorosi del pigmento superficiale. Questo adattamento cromatico può ottimizzare l'assorbimento della luce disponibile o riflettere una minore necessità di pigmenti protettivi in condizioni di luce meno intense.
Una caratteristica distintiva dei fusti sterili è la loro ramificazione. Essi presentano una dozzina di rametti, ciascuno a quattro coste, disposti in verticilli alla base delle foglie, che a loro volta si trovano nei nodi del fusto. Anche questi rametti sono articolati in nodi e relativi internodi, con quelli superiori che diventano progressivamente più brevi, conferendo alla pianta un aspetto che ricorda una piccola conifera o una "coda di cavallo". Questa struttura ramificata massimizza la superficie esposta alla luce solare, ottimizzando l'efficienza fotosintetica e supportando la crescita e il mantenimento della pianta.
Caratteristiche Comuni ai Fusti di Equisetum arvense
Nonostante le loro diverse funzioni, i fusti fertili e sterili condividono alcune caratteristiche strutturali fondamentali. Entrambi i fusti sono fortemente scanalati longitudinalmente, un tratto che li rende "alati", e sono chiaramente suddivisi in diversi nodi e relativi internodi. Le striature verticali, che possono variare da 6 a 19, presentano inoltre la particolarità di essere sfalsate passando per due internodi contigui. Questa disposizione non casuale potrebbe contribuire alla resistenza meccanica del fusto, distribuendo meglio le forze di torsione e compressione.
Internamente, i fusti di Equisetum arvense sono cavi, o fistolosi. Al loro interno, è presente una sottile cavità longitudinale centrale, conosciuta come cavità midollare. Nel caso specifico di questa specie, tale cavità è particolarmente ampia, arrivando a essere larga almeno il doppio delle cavità laterali che la circondano, e in genere 2 - 5 volte quelle laterali, occupando in tutti i casi circa 1/3 del diametro totale del fusto. Questa cavità centrale, insieme alle cavità laterali, contribuisce alla leggerezza e alla flessibilità del fusto, pur mantenendone una notevole resistenza. Inoltre, sia i fusti che i rami di Equisetum arvense sono senza vistose sporgenze silicizzate. Il diametro dei fusti in generale si aggira tra 1 e 5 mm.
Questa combinazione di caratteristiche morfologiche e anatomiche sottolinea l'efficienza strutturale ed ecologica dell'Equisetum arvense, consentendogli di prosperare in una vasta gamma di ambienti e di perpetuare la sua antica linea evolutiva.
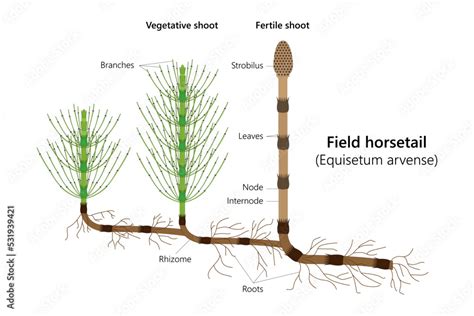
Anatomia Interna del Fusto: Una Struttura Specializzata
L'analisi di una sezione trasversale del fusto di Equisetum arvense, in corrispondenza di un internodo, rivela una notevole complessità e specializzazione strutturale, che supporta sia la funzione meccanica che quella fisiologica della pianta.
La parte più esterna del fusto è costituita da un'epidermide (e) che riveste interamente la superficie. Una caratteristica distintiva e di grande importanza di questa epidermide è la presenza di numerosi granuli di silice (biossido di silicio). Questa incorporazione di silice conferisce al fusto proprietà meccaniche notevoli, come durezza e resistenza all'abrasione e al taglio. È proprio da qui che derivano alcune delle applicazioni storiche della pianta, come l'uso per la lucidatura di oggetti.
In corrispondenza delle costole longitudinali che corrono lungo il fusto, il tessuto vegetale sottostante, chiamato cordone sclerenchimatico (s), è ulteriormente ispessito. Lo sclerenchima è un tessuto di supporto, costituito da cellule con pareti spesse e lignificate, che fornisce resistenza meccanica e rigidità alla pianta. L'ispessimento in corrispondenza delle costole aumenta la stabilità strutturale del fusto, aiutandolo a resistere alle forze esterne come il vento o la pressione dell'acqua.
Tra una costola e l'altra, si trovano delle depressioni chiamate “vallecole”, dove l'ispessimento sclerenchimatico è meno pronunciato. Nelle vallecole dei fusti sterili, è presente il parenchima clorofilliano (pc). Questo tessuto, ricco di cloroplasti, è il sito primario della fotosintesi. La sua posizione nelle vallecole, leggermente incavata, potrebbe offrire una certa protezione dagli eccessi di irradiazione solare pur mantenendo un'esposizione sufficiente. In questa stessa zona delle vallecole, si trovano anche gli stomi, o aperture stomatiche (as). La funzione degli stomi è cruciale per la vita della pianta: essi consentono lo scambio gassoso tra l'interno e l'esterno del vegetale. In particolare, attraverso gli stomi avviene la fuoriuscita di vapore acqueo, un processo noto come traspirazione, e l'entrata di anidride carbonica (CO2), essenziale per la fotosintesi, e di ossigeno (O2), necessario per la respirazione cellulare.
Più internamente, immersi nel parenchima (p) generale del fusto, si possono osservare altre strutture di grande importanza. Tra queste, spiccano i canali vallecolari (cv), ampi spazi aerei che si estendono longitudinalmente sotto le vallecole. La loro funzione non è del tutto definita, ma si ritiene che possano giocare un ruolo nel facilitare la circolazione dell'aria all'interno della pianta, un adattamento che potrebbe essere particolarmente utile per le specie che crescono in ambienti umidi o con substrati scarsamente aerati. Accanto ai canali vallecolari, sono presenti i fasci cribro-vascolari (fv). Questi sono le strutture conduttrici di sostanze liquide e nutrienti. Essi comprendono il xilema, che trasporta l'acqua e i sali minerali dalle radici alle parti aeree, e il floema, che veicola i prodotti della fotosintesi (zuccheri) dalle foglie (o in questo caso, dai fusti fotosintetici) a tutte le parti della pianta, inclusi i rizomi e i tuberi di riserva.
Questa complessa organizzazione interna del fusto dell'Equisetum arvense, con la sua combinazione di tessuti di supporto silicizzati, parenchima fotosintetico, sistemi di aerazione e vasi conduttori, è un esempio eccellente di come la natura abbia evoluto soluzioni efficienti per la sopravvivenza e la prosperità di questa pianta antichissima. Ogni componente contribuisce in modo sinergico a garantire l'integrità strutturale e l'efficienza fisiologica necessarie per la vita in ambienti spesso competitivi.
Le Foglie: Microfille Ridotte e la Loro Funzione
Le foglie nell'Equisetum arvense, come in molte Pteridophyte, non si presentano nella forma laminare e complessa tipica delle Angiosperme. In questo caso, vengono più precisamente chiamate microfille, a sottolineare la loro struttura semplice e le dimensioni ridotte. Queste microfille sono situate in corrispondenza dei nodi del fusto, sia nei fusti fertili che in quelli sterili.
Una caratteristica peculiare delle microfille di Equisetum arvense è la loro disposizione e fusione. Sono erette e appressate al fusto stesso, conferendo un aspetto compatto ai nodi. Sono concresciute le une alle altre, ovvero saldate tra loro, a formare una sorta di collaretto lobato o una guaina distintiva che circonda il fusto. Questa guaina non è differenziata in picciolo e lamina fogliare, mancano quindi le strutture tipiche delle foglie delle piante superiori. Le loro dimensioni sono tali per cui ricoprono meno della metà dell'internodo sottostante, rendendole piuttosto poco appariscenti rispetto al fusto stesso.
La forma di queste microfille è lanceolata e squamiforme, ricordando delle piccole scaglie. Ciascuna microfilla possiede un unico nervo dorsale, elemento conduttore e di supporto, e un apice acuminato, spesso di colore bruno. Il numero di denti presenti sul margine superiore di questa guaina fogliare varia solitamente tra 8 e 12. Questi denti rappresentano le punte libere delle singole microfille saldate nella parte basale, rendendo evidente la loro origine da foglie individuali.
Nei fusti sterili, dove la fotosintesi è la funzione predominante, le foglie sono così poco significative dal punto di vista della superficie fotosintetica che il fusto stesso si sostituisce ad esse per il processo fotosintetico. Questo avviene tramite degli stomi superficiali, ovvero delle cellule clorofilliane che sono a contatto con l'epidermide per lunghi tratti. Pertanto, la funzione primaria delle microfille in Equisetum arvense non è la fotosintesi, ma piuttosto la protezione dei giovani tessuti meristematici presenti nei nodi e forse, in qualche misura, il supporto strutturale del fusto. Le immagini delle foglie di un fusto fertile e di un fusto sterile, come quelle raccolte a Le Laste, Limana (BL), a 661 m s.l.m., evidenziano queste sottili differenze e la loro integrazione con la struttura generale del fusto.

Riproduzione: Lo Strobilo e la Disseminazione delle Spore
Il fusto fertile dell'Equisetum arvense è intrinsecamente legato alla riproduzione attraverso una struttura altamente specializzata posta alla sua sommità: lo strobilo. Questo apparato riproduttivo apicale è il fulcro della propagazione della specie e rappresenta l'organo che, in assenza di fiori e semi, assicura la continuità genetica.
Lo strobilo è una struttura compatta, di forma cilindrico-ovoidale, la cui lunghezza varia tipicamente tra 1,5 e 4 cm. È ricoperto quasi completamente da foglie modificate, chiamate sporofilli, che hanno una forma molto particolare: peltata. Questa morfologia significa che un corto peduncolo è inserito al centro della pagina inferiore di questa foglia modificata. La parte opposta del peduncolo si collega all'asse centrale del fusto e, di conseguenza, allo strobilo, sostenendo lo sporofillo in una posizione che ottimizza la dispersione delle spore. La forma della lamina dello sporofillo è irregolarmente esagonale, permettendo un incastro efficace con gli sporofilli adiacenti per formare la struttura compatta dello strobilo.
La vera meraviglia riproduttiva si trova al di sotto di ciascuno sporofillo. Tutto intorno all'estremità inferiore della foglia peltata, sono inseriti da 5 a 12 sporangi. Gli sporangi sono i veri e propri contenitori delle spore, le microscopiche unità riproduttive dell'Equiseto. All'interno di questi sporangi, attraverso un processo di meiosi, vengono prodotte le spore.
Le spore dell'Equisetum arvense sono del tipo isospore, il che significa che sono tutte uguali in termini di dimensione e morfologia, e non sono differenziate sessualmente a questo stadio. La loro superficie è stratificata in quattro livelli sovrapposti, ognuno con una funzione specifica. Il più importante di tutti è il primo livello, quello più esterno, chiamato esosporio. Questa membrana esterna non è una semplice protezione, ma una struttura dinamica. Lacerandosi al momento della maturazione, l'esosporio lascia libere quattro appendici nastriformi e igroscopiche chiamate "apteri". Questi apteri sono simili agli “elateri” che si trovano nelle Epatiche, un altro gruppo di piante primitive. La funzione cruciale degli apteri è quella di far muovere la spora. Essi sono dotati di movimenti igroscopici, il che significa che si aprono e si chiudono in risposta alle variazioni dell'umidità atmosferica. Quando l'aria è secca, gli apteri si dispiegano, aumentando la superficie della spora e permettendole di essere facilmente trasportata dal vento anche per lunghe distanze. Quando l'umidità aumenta, si riavvolgono, favorendo l'adesione della spora al substrato e la sua germinazione. Questo meccanismo dinamico è estremamente utile nel processo di disseminazione, garantendo che le spore vengano rilasciate e trasportate efficacemente nell'ambiente circostante per trovare un luogo adatto alla germinazione.
Il periodo di maturazione degli strobili è precoce nel ciclo vegetativo della pianta. Lo sviluppo degli strobili avviene tipicamente tra febbraio e marzo, quando la temperatura inizia a salire e l'umidità è ancora abbondante. Le spore, invece, raggiungono la piena maturazione e vengono rilasciate nel periodo di marzo-aprile. Questo tempismo è strategico: consente alle spore di essere disperse prima che la fitta crescita dei fusti sterili e di altre piante crei una barriera fisica alla loro dispersione e prima che le condizioni di siccità estiva possano compromettere la germinazione. Località come Le Laste, Limana (BL), a 661 m s.l.m., offrono ottimi esempi per osservare sia lo strobilo maturo che gli sporofilli e sporangi in dettaglio.

Spore movement of horsetails
Habitat Naturale e Distribuzione Ecologica
L'appellativo specifico “arvense” non è solo una curiosità etimologica, ma un chiaro indicatore dell'habitat preferenziale di questa specie. Esso denota la sua presenza in habitat campestri, ed effettivamente non è infrequente trovare l'Equisetum arvense prosperare su terreni incolti umidi o lungo i fossi. Questa preferenza per ambienti che presentano una buona disponibilità idrica è una caratteristica chiave della sua ecologia.
Tuttavia, la sua capacità di adattamento va ben oltre i semplici campi umidi. È una pianta estremamente versatile e può essere rinvenuta anche lungo le scarpate, che offrono spesso terreni smossi e buona esposizione, in ambienti ruderali, ovvero aree disturbate dall'attività umana come bordi stradali, macerie o terreni di risulta, e su terreni sabbiosi e argillosi. Questa ampia tolleranza a diverse tipologie di suolo e a vari gradi di disturbo ambientale testimonia la robustezza e la resilienza di Equisetum arvense. La sua diffusione è favorita dalla rete rizomatosa sotterranea, che le permette di colonizzare rapidamente nuove aree e di formare dense popolazioni, rendendola una specie a volte considerata infestante in contesti agricoli. La sua presenza è un buon indicatore di suoli con un buon drenaggio ma con una costante umidità di fondo, elementi cruciali per il suo sviluppo.
Diversità Morfologica e Ibridi: Varietà e Forme di Equisetum arvense
Nonostante la sua apparente omogeneità, l'Equisetum arvense presenta una certa diversità morfologica che ha portato alla descrizione di diverse forme (fo.), varietà (var.) e sottospecie (subsp.). Queste differenziazioni riflettono adattamenti locali o variazioni genetiche all'interno della specie, sebbene la loro classificazione possa essere oggetto di revisioni botaniche. Tra le forme descritte si trovano Equisetum arvense L. fo. pseudo-alpestre Vict. e Equisetum arvense L. fo. pseudo-varium Vict. A livello di sottospecie, è stata identificata Equisetum arvense L. subsp. boreale Á. Le varietà includono Equisetum arvense L. var. alpestre Wahlenb., Equisetum arvense L. var. nemorosum A. Br. e Equisetum arvense L. var. serotinum G.L. Mey. Ognuna di queste entità può presentare sottili differenze nella ramificazione, nella colorazione o nella taglia, spesso in relazione a specifiche condizioni ambientali di crescita.
Oltre alle variazioni intraspecifiche, l'Equisetum arvense è noto anche per la sua capacità di formare ibridi con altre specie di Equisetum. Un esempio significativo è Equisetum ×litorale Kühlewein (1845), comunemente noto come Equiseto litorale. Questo ibrido si origina dall'incrocio tra Equisetum arvense e Equisetum fluviale. Le caratteristiche di Equisetum ×litorale sono abbastanza intermedie tra le due specie di origine, manifestando un mix di tratti parentali. Una peculiarità che lo distingue è la cavità centrale dei fusti, che è molto allargata e può occupare fino ai 4/5 del diametro totale del fusto, una dimensione notevole rispetto a quella dell'Equisetum arvense puro. Questo ibrido frequenta i luoghi umidi, spesso in prossimità di corpi d'acqua, ereditando la preferenza per l'acqua di Equisetum fluviale. Tuttavia, è considerato molto raro, principalmente a causa della scarsa fertilità delle sue spore, che sono generalmente abortive. Questa limitata capacità riproduttiva sessuale è un tratto comune negli ibridi interspecifici e contribuisce alla loro rarità in natura. Un altro ibrido menzionato è Equisetum ×rothmaleri C., che, sebbene non dettagliato qui, indica ulteriormente la propensione del genere a fenomeni di ibridazione, che possono portare a nuove forme e adattamenti. Lo studio di queste varietà e ibridi è fondamentale per comprendere la dinamica evolutiva e la plasticità genetica del genere Equisetum.
Usi e Proprietà Tradizionali e Fitoterapiche
L'Equisetum arvense è una pianta che, per la sua composizione chimica e le sue proprietà fisiche, ha trovato un'ampia gamma di applicazioni nel corso dei secoli, sia in ambito tradizionale che in quello fitoterapico e domestico.
Usi Fitoterapici e Medicinali
A scopi fitoterapici, la parte della pianta tradizionalmente utilizzata è il fusto sterile. Questo viene raccolto in estate, il periodo in cui le sue proprietà sono massimamente espresse, quindi essiccato, triturato e infine polverizzato per l'uso. Le proprietà attribuite all'equiseto sono molteplici e variano dalla capacità antiemorragica e cicatrizzante, utile per favorire la guarigione di ferite e arrestare piccole emorragie, a quella emostatica. È anche riconosciuto per le sue virtù diuretiche, che lo rendono utile nel favorire l'eliminazione dei liquidi in eccesso, e astringenti. In passato, è stato impiegato anche come antitubercolare.
Tuttavia, la proprietà per cui l'equiseto è maggiormente apprezzato e studiato è quella remineralizzante. Questa caratteristica è dovuta all'elevato contenuto di acido silicico e altri minerali presenti nei fusti. Studi clinici hanno dimostrato che l'assunzione di equiseto in polvere abbrevia sensibilmente il tempo di guarigione delle fratture ossee, suggerendo un ruolo nel rafforzamento e nella riparazione del tessuto osseo. Questo lo rende un potenziale alleato nel mantenimento della salute di ossa, unghie e capelli, e nel recupero da traumi scheletrici. È importante sottolineare che le informazioni riportate non sono consigli medici e potrebbero non essere accurate; l'uso di qualsiasi rimedio fitoterapico dovrebbe avvenire sotto la guida di un professionista sanitario qualificato.
Usi Alimentari Tradizionali
In passato, presso le famiglie contadine, l'equiseto trovava anche un posto nella dieta, seppur occasionale. I giovani germogli, ancora teneri, venivano a volte impanati e fritti, o semplicemente conditi con aceto, come una sorta di verdura selvatica. Inoltre, poteva essere aggiunto a zuppe o minestroni come un integratore naturale di sali minerali, sfruttando la sua ricchezza in micronutrienti essenziali per la dieta. Questa pratica evidenzia la conoscenza empirica delle sue proprietà nutritive da parte delle popolazioni rurali.
Usi Domestici e Industriali
Le notevoli proprietà abrasive dell'acido silicico presente negli equiseti hanno portato a un uso pratico e ingegnoso della pianta, che veniva anche chiamata “erba dello stagno”. I fusti venivano sfruttati nella lucidatura di oggetti in legno o metallo: strofinandoli direttamente sulle superfici, la silice agiva come un delicato abrasivo, rimuovendo impurità e conferendo lucentezza. L'operazione risultava particolarmente pratica anche per la forma e l'elasticità dei fusti stessi, che si adattavano bene a diverse superfici. Per questa ragione, erano sovente adoperati anche per la pulizia dell'interno di vasi e bottiglie, dove la loro flessibilità permetteva di raggiungere angoli difficili.
Usi Cosmetici
Gli antichi romani, con la loro attenzione per l'igiene e la cura del corpo, utilizzavano l'Equiseto come sostituto del sapone. La sua texture abrasiva e forse le sue proprietà detergenti naturali lo rendevano adatto a tale scopo. Anche oggi, l'equiseto ha trovato applicazione in cosmetica. Entra infatti negli ingredienti di alcune creme antirughe. Si ipotizza che i suoi componenti, in particolare la silice e altri minerali, possano contribuire a rallentare l'invecchiamento della pelle, migliorando l'elasticità e la tonicità dei tessuti cutanei, suggerendo un potenziale nel mantenimento di un aspetto giovanile della pelle.

Cenni Storici ed Evolutivi: Una Pianta Fossile Vivente
L'Equisetum arvense non è solo una pianta con notevoli proprietà e un ciclo vitale interessante; è anche una testimonianza vivente di un'era geologica lontana. Si pensa che sia comparsa circa 300 milioni di anni fa, un'affermazione che la colloca tra le specie vegetali più antiche ancora presenti sul pianeta. Questa straordinaria longevità evolutiva la rende un vero e proprio "fossile vivente".
Durante il Carbonifero, il periodo a cui risale la sua comparsa, gli antenati dell'Equisetum, come i Calamiti, dominavano vaste foreste paludose, raggiungendo dimensioni arboree impressionanti, a differenza delle attuali specie erbacee. Queste foreste hanno contribuito in modo significativo alla formazione dei grandi giacimenti di carbone che oggi utilizziamo. La struttura segmentata del fusto, la riproduzione tramite spore e la presenza di silice nell'epidermide sono tutte caratteristiche che si sono conservate da questi antichi progenitori, dimostrando la straordinaria efficacia e adattabilità di questo piano corporeo vegetale.
Studiare l'Equisetum arvense significa quindi guardare non solo a una specie vegetale del presente, ma anche a una finestra sul passato profondo della vita vegetale sulla Terra. La sua persistenza attraverso ere geologiche caratterizzate da profondi cambiamenti climatici e da grandi eventi di estinzione è una prova della sua resilienza e della sua capacità di adattamento, rendendola un soggetto di studio affascinante per botanici ed evoluzionisti.