L'embriogenesi, il complesso processo che porta alla formazione di un organismo pluricellulare a partire da una singola cellula, è una danza finemente coreografata di eventi cellulari e molecolari. Tra le prime e più fondamentali strutture che emergono in questo percorso c'è il sacco vitellino, una formazione anatomica vitale nei cordati, deputata all'assolvimento delle funzioni nutrizionali nei primi stadi di sviluppo. Questo annesso embrionale, essenziale per il supporto metabolico e lo sviluppo iniziale di numerosi sistemi organici, è rivestito internamente da un foglietto embrionale primario: l'endoderma. La comprensione dello sviluppo e delle molteplici funzioni di questo endoderma all'interno del sacco vitellino e delle sue derivazioni successive è cruciale per apprezzare la complessità della vita embrionale precoce.
Il sacco vitellino è il primo elemento visibile nel sacco gestazionale durante la gravidanza, di solito intorno alle 5 settimane di gestazione, posizionandosi sulla parte ventrale dell'embrione. La sua struttura è intrinsecamente legata all'endoderma, che ne costituisce il rivestimento interno, mentre all'esterno si trova uno strato di mesoderma. Questa disposizione bilaminare è fondamentale per le interazioni che avverranno nelle fasi successive dello sviluppo embrionale, con il mesoderma e l'ectoderma che si formano durante la gastrulazione nella terza settimana di sviluppo. La gastrulazione, in effetti, è un processo che include una transizione epitelio-mesenchimale primaria, ponendo le basi per la formazione di tutti e tre i foglietti embrionali.
Origine e Prime Fasi di Sviluppo del Sacco Vitellino
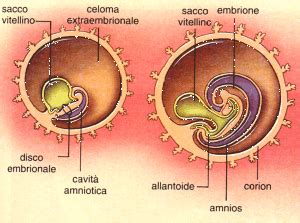
Il sacco vitellino inizia a formarsi durante la seconda settimana dello sviluppo embrionale, contestualmente alla costituzione della vescicola amniotica. In questa fase precoce, la blastocisti, accresciutasi, tende ad uscire dalla mucosa uterina occupando il lume dell'utero, un processo fondamentale per l'impianto embrionale che caratterizza la seconda settimana. Durante questo periodo, si osserva anche il differenziamento del trofoblasto in citotrofoblasto e sinciziotrofoblasto, con la formazione del sistema lacunare, tutti elementi che preparano il terreno per gli scambi con l'ambiente materno. Contemporaneamente, si assiste alla formazione del mesoderma extraembrionale e della cavità corionica, o celomatica extraembrionale, che circonda l'embrione e i suoi annessi.
La trasformazione del sacco vitellino da primario a secondario è un momento chiave nello sviluppo. Questa prima modificazione è determinata dalla delaminazione del magma reticolato, noto anche come reticolo extraembrionale, che riveste esternamente il sacco. Questo evento morfogenetico porta a una strozzatura nella sua parte terminale, specificamente nella zona di connessione tra l'ipoblasto e la membrana esocelomica di Heuser. Le due parti risultanti si distaccano, e quella inferiore, di dimensioni minori, forma una cisti che verrà presto riassorbita, lasciando il sacco vitellino secondario come struttura dominante.
Nel punto di giunzione fra endoderma e membrana esocelomica, il mesoderma extraembrionale si accresce, comprimendo il sacco vitellino sino a che la membrana esocelomica viene staccata, si raggrinzisce e scompare all'interno del mesoderma. Il mesoderma, in tale momento, si lamina creando delle cavità che si uniscono tra di loro, formando un'unica grande cavità. Il foglietto più esterno di questa cavità, denominato parietale, si assembla al citotrofoblasto, portando alla creazione di un altro annesso embrionale, il corion, che andrà a costituire, assieme alla mucosa uterina, la placenta. Contestualmente, il foglietto viscerale più interno si lega all'endoderma, modificando il sacco vitellino primario che, a questo punto, diverrà di tipo secondario, caratterizzato da due pareti. Questa riorganizzazione conferisce al sacco vitellino secondario una cruciale capacità: quella di generare parte dei vasi sanguigni dell'embrione. Il corion, nel frattempo, consentirà gli scambi di ossigeno tra l'embrione e la madre, evidenziando l'interconnessione degli annessi embrionali.
Il Sacco Vitellino come Centro di Ematopoiesi e Vasculogenesi Embrionale Precoce
Una delle funzioni più significative del sacco vitellino, mediata in parte dalle interazioni tra il suo endoderma e lo strato mesodermico adiacente, è il suo ruolo precoce nella formazione del sistema circolatorio e nella produzione di cellule del sangue. Il sacco vitellino, infatti, ha la capacità di generare parte dei vasi sanguigni dell'embrione, un evento fondamentale per l'instaurarsi di un sistema circolatorio funzionante.
Il sangue è convogliato alla parete del sacco vitellino dall'aorta primitiva; dopo aver circolato attraverso un plesso capillare a maglia larga, che rappresenta un abbozzo del sistema circolatorio, viene restituito dalle vene vitelline al tubo cardiaco dell'embrione. Questo circuito primitivo sottolinea l'importanza del sacco vitellino come sito iniziale di vasculogenesi e angiogenesi. Le isole angioblastiche extra-embrionali, che si formano in questa fase, sono le prime strutture deputate alla genesi dei vasi sanguigni e delle cellule ematiche.
Inoltre, il sacco vitellino è una delle sedi primarie di emopoiesi durante lo sviluppo embrionale. È qui che si originano le cellule staminali emopoietiche del sacco vitellino, che successivamente migreranno verso altre regioni, come la regione aorta-gonado-mesonefrica, e poi in altre sedi di emopoiesi prima e dopo la nascita. Queste cellule sono essenziali per la formazione di tutti i tipi di cellule del sangue. Questo ruolo è transitorio ma assolutamente critico, in quanto getta le basi per la futura produzione di sangue nell'organismo in sviluppo. L'espansione del sacco vitellino e la sua ricca vascolarizzazione sono indicatori della sua attività metabolica e ematopoietica in questo stadio.
Embriologia 03 - Seconda, terza, quarta settimana e organogenesi
Il Sacco Vitellino Definitivo e l'Organogenesi dell'Intestino
La quarta settimana di sviluppo è un periodo di intense modificazioni morfologiche, caratterizzate in particolare dal ripiegamento dell'embrione. Durante questa settimana, avviene la determinazione delle superfici embrionali, e il sacco vitellino subisce esternamente la pressione esercitata dalle pieghe embrionali: la piega cefalica, la piega caudale e le pieghe laterali. Questo processo di ripiegamento laterale e cefalo-caudale è cruciale e porta alla definizione cilindrica del corpo embrionale, trasformando una struttura laminare in una forma tridimensionale.
Come conseguenza di queste dinamiche di ripiegamento, il sacco vitellino non rimane una struttura completamente separata, ma una piccola porzione di esso, situata nella parte superiore, viene incorporata all'interno del corpo embrionale, andando a costituire il tubo intestinale. La parte immediatamente inferiore a questa porzione incorporata, invece, forma il mesentere ventrale, una piega peritoneale che supporta e vascolarizza l'intestino. Parallelamente, l'ammios, che continua ad espandersi velocemente, giunge a "risucchiare" parte del sacco vitellino secondario. Questa porzione assorbita dall'amnios costituirà l'intestino medio, un segmento fondamentale del canale alimentare. L'intestino medio non si richiude subito, poiché rimane in comunicazione con il vitello o, in una fase successiva, con il cordone ombelicale attraverso il dotto vitellino. Questa connessione persiste per un certo periodo, riflettendo la sua origine dal sacco vitellino.
L'Endoderma: Origine e Differenziamento dell'Apparato Digerente e Respiratorio
L'endoderma è, per definizione, il foglietto embrionale da cui si origina l'intestino, sia anteriore che posteriore, una volta che il tubo digerente primitivo si è completamente richiuso all'interno dell'embrione. Quando l'embrione è ancora laminare, il canale alimentare è composto da tre parti distinte: cefalica, media e claucale. Le aree cefalica e claucale, prima della nascita, sono chiuse. Il processo di organogenesi, che porta allo sviluppo degli abbozzi di organi, è fortemente guidato da interazioni e induzioni tra i diversi foglietti embrionali.
L'Intestino Anteriore e i Suoi Derivati Endodermici
La regione craniale del tubo digerente, rispetto al punto d’origine dell’albero respiratorio, è definita faringe. Da essa si originano le strutture del faringe branchiale, che in tutti i vertebrati si espande in modo segmentato fino a formare 6 coppie di archi branchiali. In corrispondenza delle estroflessioni degli archi, si formano delle introflessioni, le prime dette solchi branchiali e le seconde tasche branchiali. Tra un solco e una tasca è presente del mesoderma, e sebbene negli amnioti la fessurizzazione di queste strutture non avvenga come negli organismi acquatici, le loro derivazioni sono cruciali.
Negli organismi polmonati, il primo abbozzo dell'apparato respiratorio avviene per evaginazione del faringe in una zona più posteriore. L'albero respiratorio, infatti, si forma per evaginazione del tubo digerente nei rettili, uccelli e mammiferi, evidenziando una differente origine rispetto alle branchie. Nel pavimento del faringe, che è parte dell'intestino cefalico, si forma una estroflessione con un diverticolo che si differenzierà nella "Doccia laringotracheale", segnando l'origine del sistema respiratorio. La doccia poi si biforca per formare le gemme polmonari, e all'estremità dei bronchioli si avranno estroflessioni sacciformi che formeranno gli alveoli, le unità funzionali dello scambio gassoso. Inizialmente, il diverticolo è attaccato al faringe, ma successivamente si distaccherà, diventando una struttura indipendente, seppur con un'origine comune.
Posteriormente al faringe, il tubo digerente si restringe per formare l'esofago, e poi si dilata a formare lo stomaco. L'esofago presenta muscolatura liscia che consente contrazioni peristaltiche, fondamentali per far scorrere il bolo alimentare. L'epitelio che riveste lo stomaco possiede ghiandole che producono acido cloridrico (HCl), mantenendo un pH interno di circa 4.5, e ghiandole che producono pepsinogeno, che proprio in ambiente acido si trasforma in pepsina, la sua forma attiva e digestiva.
Dalla stessa evaginazione del tubo digerente primitivo, o da essa gemmano, fegato, cistifellea e pancreas. Nella parete ventrale si forma un diverticolo epatico, e da questo fegato primitivo si formerà poi il diverticolo della cistifellea. La proliferazione primaria delle cellule endodermiche forma il parenchima epatico, mentre la proliferazione secondaria origina la cistifellea. Prima dell'intestino tenue, e in particolare nel duodeno, trovano sbocco i dotti pancreatici e i dotti biliari provenienti dal fegato e dalla cistifellea, tutti derivati endodermici cruciali per la digestione. La milza, pur essendo adiacente, è di origine mesodermica e non endodermica. La formazione della cavità pericardica, pleurica e peritoneale, così come lo sviluppo del diaframma, sono processi complessi che coinvolgono anche i derivati endodermici nella loro modellazione.
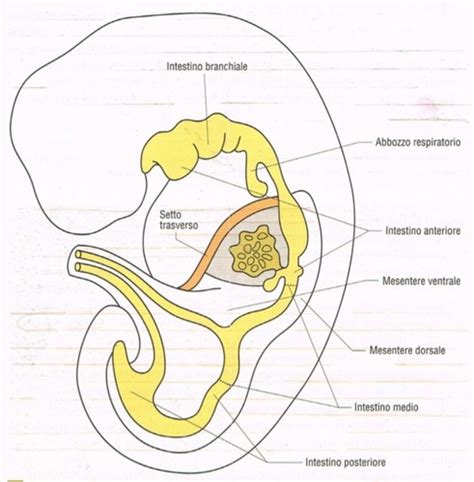
L'Intestino Medio e Posteriore
L'intestino medio è la porzione da cui si sviluppano l'intestino tenue, il cieco, il colon ascendente e i due terzi del colon trasverso. Questo segmento subisce importanti rotazioni durante lo sviluppo embrionale, un processo essenziale per la sua corretta posizione nell'addome e per la sua vascolarizzazione.
L'intestino posteriore primitivo, invece, dà origine all'ultimo terzo del colon trasverso, al colon discendente, al colon sigmoide, al retto e alla parte superiore del canale anale. La cloaca e la sua sepimentazione sono caratteristiche distintive di questa regione. Da qui, inoltre, si sviluppano la vescica e l'uretra, entrambe di derivazione endodermica. La formazione dell'apparato urogenitale è un processo complesso che coinvolge interazioni genetiche e ormonali, con l'endoderma che contribuisce specificamente alla vescica e all'uretra. Il trigono vescicale, una parte della vescica, ha una derivazione mesodermica ma è strettamente integrato con le componenti endodermiche.
Interazioni Morfogenetiche e Molecolari Nello Sviluppo dell'Endoderma
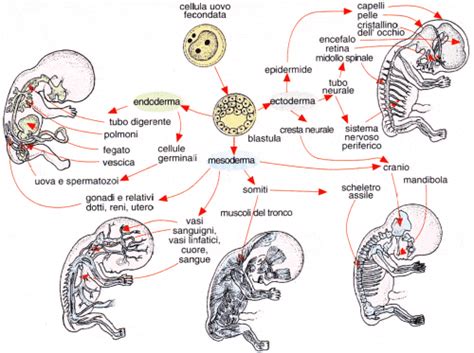
La determinazione dei foglietti embrionali, incluso l'endoderma, è un processo finemente regolato che inizia con la gastrulazione nella terza settimana di sviluppo. Durante questo periodo, l'embrione, che si espanso solo in superficie e formato da ectoderma e endoderma, si evolve. Si origina una sporgenza nella zona anteriore denominata tubercolo precordale, e si forma un solco per effetto dello scivolamento di alcune cellule dell'ectoderma che scivolano sotto le altre, moltiplicandosi sino a creare un terzo strato: il terzo foglietto embrionale o mesoderma, dal quale si svilupperanno i tessuti connettivi e muscolari. Questo mesoderma si sviluppa prevalentemente lungo i lati, originando le ali mesodermiche che si accrescono sino a saldarsi col mesoderma extraembrionale. Infine si uniscono tra di loro davanti alla placca precordale e, dal punto di unione, prenderà vita l'area cardiogenica, sede di sviluppo del cuore.
L'organogenesi, il processo che porta allo sviluppo degli abbozzi di organi, si basa su complesse induzioni reciproche tra i foglietti embrionali. Ad esempio, la notocorda (di origine mesodermica) istruisce l’ectoderma sovrastante a diventare ectoderma neurale, il resto dell’ectoderma diventerà invece epidermide. Questo processo inizia quindi per induzione primaria, in quanto una specifica regione dell’embrione interagendo con un’altra regione ne influenza il differenziamento lungo una via differenziativa. Similmente, lo sviluppo dell'endoderma e delle sue derivazioni, come fegato e pancreas, è guidato da segnali induttivi provenienti dal mesoderma e da altri tessuti circostanti.
I geni omeotici (HOX) sono fondamentali nello schema corporeo lungo l'asse antero-posteriore, influenzando la regionalizzazione dell'intestino derivato dall'endoderma e la specificazione dei diversi tratti del canale alimentare. Meccanismi molecolari complessi, come il modello "clock and wavefront" per la somitogenesi (sebbene legata al mesoderma parassiale), illustrano come processi ritmici e gradiente di fattori di crescita influenzino la formazione delle strutture corporee, inclusi i tessuti adiacenti di origine endodermica. La regressione della linea primitiva, ad esempio, è un processo importante che, se alterato, può portare a malformazioni come il teratoma sacro-coccigeo, evidenziando la sensibilità di queste prime fasi di sviluppo.
La determinazione degli assi corporei - antero-posteriore, dorso-ventrale e sinistro-destro - è cruciale e coinvolge gradienti decrescenti di fattori della famiglia del TGFbeta, come Nodal e BMP. Questi segnali molecolari influenzano direttamente o indirettamente il destino delle cellule endodermiche e la loro organizzazione spaziale. Il corretto funzionamento di questi meccanismi assicura la corretta formazione e posizionamento degli organi derivati dall'endoderma, come il fegato, il pancreas e il tratto gastrointestinale.
Connessioni del Sacco Vitellino con Altri Annessi Embrionali
Il sacco vitellino non agisce in isolamento, ma è parte integrante di un sistema di annessi embrionali che include anche la placenta, l'allantoide e il cordone ombelicale. Questi annessi lavorano in sinergia per sostenere lo sviluppo dell'embrione e del feto.
Nella porzione di magma non delaminato, si crea il peduncolo d'attacco, una struttura che diverrà in seguito il cordone ombelicale. Questa evoluzione stabilisce una connessione vitale tra l'embrione e gli annessi extraembrionali, facilitando lo scambio di nutrienti e cataboliti. Il cordone ombelicale, in effetti, contiene i vasi vitellini e successivamente i vasi ombelicali che si sviluppano per garantire questa comunicazione.
L'allantoide stessa costituisce una evaginazione del sacco vitellino che cresce e si lega al peduncolo d'attacco. Questa struttura, pur avendo un ruolo più significativo in altre specie per lo stoccaggio dei rifiuti e lo scambio di gas, nei mammiferi contribuisce alla formazione dei vasi ombelicali e della vescica. L'endoderma dell'allantoide, in particolare, è coinvolto nella formazione della vescica e dell'uretra, come menzionato in precedenza.
Il corion, sviluppato a partire dal mesoderma extraembrionale e dal citotrofoblasto, è un altro annesso fondamentale. Su tutta la sua superficie si creano una serie di sporgenze definite villi d'attacco. Intanto, la blastocisti tende ad occupare il lume dell'utero. Il corion inizia a perdere i villi in una delle sue due porzioni, che verrà definita "liscia", mentre nell'altra estremità i villi si ramificano ancora di più, originando la porzione "frondosa". Quest'ultima andrà a formare la componente fetale della placenta, cruciale per gli scambi tra madre e embrione/feto. La placenta, con la sua complessa struttura e funzione, è essenziale per la nutrizione, la respirazione e l'eliminazione dei rifiuti del feto, supportata anche dagli ormoni placentari che regolano la gravidanza. La circolazione placentare è un sistema altamente specializzato che garantisce questi scambi vitali.
In sintesi, l'endoderma del sacco vitellino e le sue successive derivazioni rappresentano un pilastro fondamentale dell'embriogenesi precoce, orchestrando non solo la nutrizione iniziale, ma anche la formazione del sistema circolatorio, del canale alimentare e di organi vitali come fegato, pancreas e apparato respiratorio. La sua interazione con gli altri foglietti embrionali e annessi è un esempio eloquente della complessità e dell'interdipendenza dei processi che portano alla formazione di un organismo completo.