Arriva un momento a scuola in cui si parla della riproduzione degli animali, un tema che svela la straordinaria complessità della vita. Istintivamente, tutti noi sappiamo di cosa stiamo parlando: la riproduzione è scritta nel nostro DNA e rientra tra i comportamenti fondamentali di un organismo. Gli sforzi riproduttivi nel regno animale possono essere faticosi, manifestandosi attraverso organi vistosi ed ingombranti come lunghe code, palchi e creste, complessi rituali di accoppiamento, pericolosi combattimenti e lunghe cure parentali. A livello evolutivo, ogni animale ha come scopo fondamentale della propria esistenza quello di riuscire a tramandare i propri geni, attraverso il concepimento della prole. Questo principio guida un'incredibile diversità di strategie, molte delle quali coinvolgono l'interazione tra più individui, talvolta un intero gruppo, nel processo che culmina nella fecondazione. In un mondo dove anche la festa di San Valentino è un momento di ostentazione di capacità seduttive per molti esemplari della nostra specie, è naturale chiedersi se queste dinamiche si replichino anche nel resto del regno animale.

Rituali di Corteggiamento e Forme Inusuali di Accoppiamento
La concorrenza per la riproduzione si manifesta in molte forme, spesso attraverso esibizioni che precedono la fecondazione. Non si tratta solo di dimostrare forza bruta; in molti casi, l'ostentazione del proprio vigore fisico è sufficiente. Il palco o il bramito del cervo, la coda del pavone, il corno o le mandibole di certi coleotteri, sono tutti apparati che servono, la maggior parte delle volte, a dissuadere il contendente dallo scontro fisico. Questo non è segno di codardia, ma di un rapido calcolo costi-benefici: è meglio lasciar perdere la contesa con chi può evidentemente “riempire di mazzate”, per cercare invece lo scontro con qualcuno alla propria portata. I rituali di accoppiamento del regno animale sono tra i comportamenti più affascinanti offerti dalla natura.
Un caso bizzarro è quello di una salamandra nordamericana, la cui fecondazione delle uova è interna, ma senza copula, come del resto accade per molti altri anfibi. Il maschio, infatti, depone una sacca spermatica (spermatofora) sul fondo dello stagno e poi inizia un lungo corteggiamento fatto di spinte sul muso, “carezze” sul collo e la schiena, per indurla a raccogliere con la propria cloaca la spermatofora. Anche in alcuni artropodi terrestri, come gli insetti che rappresentano tipi filogenetici più basali (cioè primitivi), il maschio deposita gli spermatozoi sul substrato, a volte conservati in speciali strutture, e induce col corteggiamento la femmina a introdurli nella sua apertura genitale. Non c'è penetrazione vera e propria. Nelle libellule si tratta di sterniti modificati del secondo segmento addominale, nei ragni dei pedipalpi.
I maschi di molte specie di ragno, ad esempio, per poter placare gli istinti cannibali delle proprie consorti che possono in alcuni casi essere diverse volte più grandi, distraggono le partner regalando prede incartate nella tela. I maestri nell’arte del corteggiamento sono senza dubbio gli uccelli del paradiso, una famiglia di uccelli endemica della Nuova Guinea. Code arricciate, pomposi collarini dai colori sgargianti petti iridescenti, associati a pose tali da farli assomigliare a maschere tribali, sono uno spettacolo irresistibile per le femmine, che valutano le esibizioni dei maschi con occhio attento. Questo è un buon esempio di comportamento riproduttivo elaborato. La selezione sessuale ha favorito lo sfoggio delle proprie abilità da parte dei maschi migliori: per una femmina l'investimento materno è importantissimo, e si tende a intraprenderlo concedendosi al più forte, al più bello, al più potente. In tal modo si garantiranno i geni migliori alla prole e quindi maggiori probabilità di sopravvivenza. Infine, è bene ricordare che accoppiarsi con un individuo di una specie diversa non sarebbe solo tempo sprecato, ma anche un investimento sbagliato e rischioso: i rituali servono quindi anche a ribadire chiaramente la reciproca appartenenza alla stessa specie. In zoologia, l'accoppiamento è l'incontro di due organismi di sesso opposto o ermafroditi a fini riproduttivi e, tra gli animali sociali, anche per allevare la propria prole. Per penetrazione sessuale si intende invece l'unione vera e propria degli organi genitali di due animali fertili che porti all'inseminazione e alla fecondazione. Negli esseri umani, diversamente da quanto accade nella maggior parte degli animali, la penetrazione non sempre è associata alla riproduzione; in molti casi essa avviene per puro piacere.
Il Prezzo della Riproduzione: Rituali di Seduzione e Dominio – Documentario Animali
Fecondazione: L'Incontro Essenziale dei Gameti
La fecondazione consiste nella fusione di due gameti, uno spermatozoo e una cellula uovo, ciascuno proveniente da un individuo genitore. Questa unione porta alla formazione dello zigote, la cellula diploide da cui si svilupperà il nuovo organismo. Nel punto in cui il primo spermatozoo tocca la superficie della cellula uovo, si forma immediatamente un'estroflessione del citoplasma, nota come cono di fecondazione, che ingloba lo spermatozoo e, ritirandosi, lo porta all'interno della cellula uovo. La fecondazione deve avvenire in un ambiente acquoso o umido, nel quale lo spermatozoo possa muoversi per raggiungere la cellula uovo.
La fecondazione può essere esterna o interna. Per gli esseri viventi che si trovano in ambiente acquatico, ciò non è un problema, e i gameti possono essere lasciati liberi all’esterno. Questa strategia è utilizzata ad esempio dagli Echinodermi, come ricci e stelle di mare. Tuttavia, presenta alcuni svantaggi, come la necessità di produrre una quantità molto elevata di gameti per aumentarne la probabilità di incontro. Nella fecondazione esterna, l'incontro dei gameti avviene in acqua e i gameti devono essere prodotti in grande quantità, poiché molti non riescono a incontrarsi.
Nei pesci e negli anfibi, la fecondazione esterna può avvenire tramite la deposizione dei gameti in un nido o comunque in raggruppamenti definiti. I gameti possono essere deposti in contemporanea o con uno sfasamento temporale, e non sempre le uova sono le prime ad essere rilasciate. Ad esempio, in molte specie di rana, i maschi emettono i propri spermi in un nido prima che la femmina vi deponga le uova. Nella maggior parte dei casi, è la femmina a deporre le uova e successivamente il maschio vi rilascia sopra lo sperma, i cui spermatozoi, grazie a particolari meccanismi di riconoscimento, riconosceranno i gameti femminili e li feconderanno. Molti altri animali si riproducono attraverso la fecondazione esterna, inclusi molti vertebrati basali.

Nella fecondazione interna, il gamete maschile, lo spermatozoo, viene introdotto nel corpo della femmina, dove si trova la cellula uovo, il gamete femminile. Per questo tipo di fecondazione sono necessari apparati riproduttori specializzati per l'introduzione e la conservazione degli spermatozoi. La scelta tra le due differenti modalità di fecondazione è legata al grado di svincolamento dall’acqua delle specie considerate e dal vantaggio in termini energetici che l’una o l’altra forniscono. Con la conquista delle terre emerse, si è dovuto far fronte al problema di dover fare incontrare i due gameti in un ambiente protetto dal disseccamento: l'utero femminile. Ecco che i maschi e le femmine delle varie specie terrestri hanno iniziato a cercarsi, richiamarsi ed attrarsi con segnali vocali, visivi e chimici al fine di avere un rapporto sessuale.
Gli uccelli presentano tutti fecondazione interna. In questo gruppo, le parti terminali dei canali digerente, urinario e genitale possiedono un tratto in comune, detto cloaca, che comunica con l’esterno. La fecondazione avviene per contatto delle cloache, e il maschio immette così nella femmina lo sperma. L’uovo viene fecondato, e nell’uovo di gallina ad esempio il gamete femminile è il tuorlo. In seguito scende lungo l’ovidutto della femmina. Durante il tragitto vengono aggiunte alcune sostanze accessorie come l’albume, che ha funzione nutritiva, di riserva di acqua e di protezione, e il guscio, solitamente rigido e poroso.
Dallo Zigote al Nuovo Organismo: Strategie di Sviluppo Embrionale
Il primo periodo di sviluppo embrionale è un momento estremamente delicato per gli animali. Le sostanze nutritive garantite dalla mamma sono un piatto ambito da tantissimi predatori, in certi casi anche della propria stessa specie. Ogni specie ha periodi di gestazione diversi. Per esempio, la gestazione del cane è lunga circa due mesi, un capodoglio ne impiega quattordici, e nella nostra specie è notoriamente di nove.
Dopo la fecondazione, la cellula uovo può svilupparsi nel corpo materno, come nel caso dei mammiferi e delle specie vivipare e ovovivipare, oppure essere deposta circondata da una struttura di protezione più o meno consistente, come negli uccelli.
I rettili e gli uccelli sono perciò ovipari, poiché le loro uova vengono depositate nell’ambiente e contenute internamente, o protette da un guscio calcareo che si forma dopo la fecondazione. Questo guscio permette all’embrione di nutrirsi sino alla schiusa. Il numero di uova prodotte è relativamente piccolo in queste specie. I mammiferi monotremi, come l’ornitorinco e l’echidna, sono anch'essi ovipari.
Esistono specie, però, che si trovano in una situazione intermedia, definite ovovivipare. Attenzione però: specie vivipare e ovovivipare possono confondersi facilmente! La vipera è un rettile che deposita le sue uova nell’ambiente solo quando lo sviluppo degli embrioni è completato. In questo caso, le uova si rompono e ne escono i piccoli perfettamente formati.
Infine, ci sono le specie vivipare, in cui lo sviluppo dell'embrione avviene interamente nel corpo materno, in un organo chiamato utero. Queste specie possiedono una placenta, un organo con vasi sanguigni che permette lo scambio di sostanze nutritizie e di rifiuto tra madre ed embrione. Lo sviluppo avviene per un periodo detto gestazione. I mammiferi non sono gli unici animali vivipari.
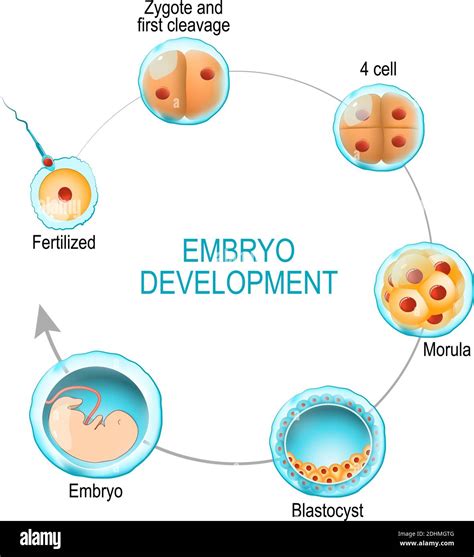
Diversità Riproduttiva e Adattamenti Specializzati
La biodiversità di forme e colori del regno animale è accompagnata da una grande diversità di comportamenti ed abitudini anche in campo sessuale. Noi umani siamo abituati ad una stretta differenziazione individuale per quanto riguarda il sesso biologico. Molte specie, però, possono cambiare il proprio sesso durante lo sviluppo, come alcuni pesci, ad esempio i pesci pagliaccio, dove un "grande Nemo" diventerà una femminuccia. Altre specie possono essere contemporaneamente maschi e femmine allo stesso momento; in tal caso si parlerà di specie ermafrodita. Le lumache di terra, lente per antonomasia, hanno grandi vantaggi dall’essere sia maschi che femmine: qualsiasi individuo incontrino della propria specie è un possibile partner.
Un altro comportamento riproduttivo estremamente affascinante è quello attuato da tantissimi insetti ed alcuni rettili, e anche uccelli secondo studi recenti, in cui la femmina può riprodursi anche senza la fecondazione maschile. Questo processo è chiamato partenogenesi, ed è molto utile in popolazioni in cui l'incontro con un maschio può risultare complicato.
Quasi tutti gli organismi viventi si affidano in parte o totalmente alla riproduzione sessuata, una strategia riproduttiva che prevede la produzione di speciali cellule dette gameti e il successivo incontro ed unione con le corrispettive cellule di un partner, rigorosamente della stessa specie, formando una nuova cellula detta embrione. Un meccanismo più semplice, invece, è detto riproduzione asessuata e consiste nel produrre organismi geneticamente identici a sé stessi senza l'aiuto di nessuno. Come in tutte le strategie vitali, vi sono vantaggi e svantaggi in entrambe le modalità. La riproduzione sessuata è preferita per una questione di ricombinazione genetica, un modo per "rimescolare le carte" garantendo una maggiore adattabilità della prole e quindi maggiori possibilità di sopravvivenza a lungo termine. Le implicazioni evolutive di queste due strategie riproduttive sono sorprendenti. Ad esempio, l'evoluzione ha favorito il differenziarsi di due sessi all'interno di ogni specie: l'incontro dei due gameti è facilitato se uno dei due, lo spermatozoo, leggero e veloce, si sposta nuotando alla ricerca di un secondo, l'ovulo, ricco di sostanze nutritive ma statico. Il "ventre" primordiale, luogo dei primi incontri amorosi, è l'Oceano, essendo il nostro regno originario dei mari paleozoici. Le specie acquatiche molto spesso si limitano ad emettere i gameti, affidandosi alle correnti per la fecondazione.
La Fecondazione di Gruppo e l'Organizzazione Sociale
La fecondazione di gruppo non si limita solo a eventi di massa come gli ammassi cotonosi e gelatinosi, ma si estende a complesse interazioni sociali che influenzano le strategie riproduttive all'interno di una comunità. Queste dinamiche sono più complesse e si sono evolute da forme che erano in principio solitarie, ponendo questioni apparentemente irrisolvibili riguardo al successo riproduttivo differenziato dell'individuo. La presenza di una forte territorialità è un fattore chiave in molti di questi sistemi, e il territorio è solitamente quello dei propri genitori. Alcuni animali si riuniscono amichevolmente in un dato territorio, mentre altri lo difendono ferocemente.

Un caso bizzarro è quello della Passera scopaiola (Prunella modularis), un simpatico uccelletto che vive ai margini delle zone boscate alpine, il cui nome è tutto un programma. Per questa specie, la dimensione del territorio gestito dal maschio influenza la strategia riproduttiva: può essere monogamica, se il territorio offre risorse sufficienti per la sola coppia; poliginica, se un maschio potrà difendere un territorio abbastanza grande da poter sostenere due femmine; oppure poliandrica, se un solo maschio non è in grado di sorvegliare adeguatamente il territorio, tanto che un maschio secondario può entrarvi e raggiungere la femmina, solitamente ben disposta verso gli intrusi. In questo scenario, a rimetterci sarà il maschio principale, che vedrà diminuito il proprio successo riproduttivo, oltre a dover gestire un territorio troppo grande per lui: si può dire quindi “cornuto e mazziato”.
Le gerarchie sociali influenzano profondamente la riproduzione in molti gruppi animali. Un esempio classico è la "scala di beccata" dei polli. Una gallina convive per un certo periodo di tempo con le altre galline senza essere beccata a sua volta, pur non poterne beccare alcuna. Una gallina dominante è solitamente ben curata e ha un andamento fiero, mentre gli individui sottomessi tendono a rimanere ai margini del gruppo. Questo schema illustra la sottomissione di una gallina rispetto a un'altra, una dinamica complessa ai fini della sopravvivenza e della riproduzione tra individui della stessa specie.
L'eusocialità rappresenta una delle forme più estreme di riproduzione di gruppo, tipica degli insetti eusociali. Prendiamo l'esempio delle api. Una colonia tipicamente è composta da 30.000-40.000 operaie e una regina adulta. La regina è l'unica femmina fertile e può deporre fino a 2000 uova al giorno, fecondate dagli spermatozoi durante l’accoppiamento. La maggior parte delle uova darà origine a femmine operaie, ma alcune svilupperanno i fuchi, che sono aploidi. Una larva di regina si sviluppa da una singola cella di cera da un uovo deposto dalla regina. Le operaie ricoprono la cella di cera, sigillandola. Quando la colonia necessita di una nuova regina, le operaie creano speciali "celle reali" contenenti cibo abbondante per la larva, che la sviluppa in una regina fertile, invece che in un'operaia sterile. La prima larva di regina che nasce uccide tutte le altre "candidate". Le operaie, pur sterili, contribuiscono indirettamente alla riproduzione del gruppo svolgendo una serie di impieghi successivi: si nutrono con il nettare e il polline raccolti per le prime tre settimane, si occupano dell'accumulo di cibo per la regina, i fuchi e le larve, producono cera usata per ingrandire il favo, e in seguito si dedicano alla raccolta di nettare e polline nell'ambiente circostante. Un feromone, prodotto dalla regina, inibisce la maturazione sessuale delle operaie, mantenendo l'equilibrio riproduttivo della colonia. Le operaie producono miele in quantità tale da soddisfare le reali necessità della colonia, ma che spesso è superiore al normale. Questo comportamento collettivo e cooperativo assicura la sopravvivenza e la riproduzione del gruppo nel suo insieme.
Un altro aspetto fondamentale della riproduzione di gruppo è l'altruismo, specialmente tra individui imparentati. Il successo riproduttivo può agire non solo sui singoli ma sulle famiglie, in cui individui imparentati si riproducono con tassi differenti. Questo è il concetto di selezione di parentela, dove il successo riproduttivo di un individuo può essere misurato anche dalla sopravvivenza dei suoi parenti, che condividono parte dei suoi geni. Ad esempio, il sistema sociale del dingo, che è una coppia monogama, dei figli, e uno-tre fratelli, mostra come gli aiutanti alla distribuzione del cibo e provvedono alla loro toeletta. È stato osservato che, con tre aiutanti, ne sopravvivevano in media sei di più. Questo dimostra come la cooperazione e il supporto all’interno di un gruppo familiare possano aumentare il successo riproduttivo complessivo. Gli organismi possono essere visti come "strumenti" ed eliminando gli alleli concorrenti.
La Consanguineità (Inbreeding) e le Sue Implicazioni nelle Popolazioni
La consanguineità, o inbreeding, è un fenomeno cruciale nelle dinamiche riproduttive di gruppo, specialmente nelle popolazioni isolate o sottoposte a selezione intensiva. Secondo la definizione di Falconer (1996), per consanguineità si intende l’accoppiamento fra individui che sono imparentati fra loro grazie ad antenati in comune. In tale situazione, il maschio e la femmina, con copie di uno stesso gene derivante dall’antenato comune, che si accoppino, possono passare tali copie alla discendenza, che sarà formata quindi da individui consanguinei. Il coefficiente di consanguineità di un soggetto (F) è la probabilità che due alleli in un locus siano identici per discendenza, ovvero provengano da un antenato comune al padre ed alla madre. Il coefficiente di consanguineità si riferisce sempre al singolo individuo e descrive il grado di parentela, che al contrario di F si riferisce a coppie di individui, fra i suoi genitori, pari a 2*F.
Esistono casi in cui la consanguineità è stata, ed è utilizzata, intenzionalmente e miratamente, ad esempio per ottenere animali da laboratorio geneticamente omogenei o per produrre, sia nel caso dei vegetali che nel caso di animali, animali altamente omozigoti da accoppiare fra loro per ottenere una progenie ibrida con performance maggiori di quelle dei genitori. In altri casi, la consanguineità è generata non intenzionalmente ma è il risultato di una situazione particolare, negli animali, ad esempio, nelle popolazioni a limitato numero di soggetti o nel caso di selezione.
Nelle moderne tecniche di allevamento, specie per i bovini da latte, l’attività di selezione è arrivata ad una grandissima efficienza, aiutata dalla possibilità di selezionare pochi animali di altissimo valore genetico con un'accuratezza di stima sempre maggiore. Negli animali sottoposti a schemi di selezione, buona parte delle razze possono essere considerate, dal punto di vista della genetica di popolazione, come una popolazione di limitata consistenza in quanto esiste un contributo sbilanciato di un limitato numero di riproduttori dovuto alla selezione che, oltre tutto, comporta un alto rischio di scegliere riproduttori imparentati fra loro. Conseguenza naturale di quanto detto è la nascita di animali sempre più consanguinei.
Tale fenomeno, ben noto agli allevatori e studiato a livello di ricerca, è dovuto a basi genetiche, ma ancora non si ha una definizione certa di esse; esistono tuttavia tre teorie che cercano di spiegare il fenomeno. Secondo la prima teoria, detta “ipotesi di dominanza parziale”, la consanguineità aumenta l’omozigosi e quindi l’espressione di alleli omozigoti recessivi con effetti fenotipici negativi. La seconda teoria, detta “ipotesi di superdominanza”, suppone che gli eterozigoti siano superiori agli omozigoti, sia dominanti che recessivi, per cui l’aumento di omozigosi causa una diminuzione dell’espressione dei fenotipi derivanti da eterozigoti.

Una volta ricavato il coefficiente F per ogni animale, è possibile quantificare l’effetto della depressione da consanguineità. Il metodo classico di calcolo è stimare l’effetto come coefficiente di regressione del fenotipo del carattere sul coefficiente F individuale. Si è visto, tuttavia, che se F viene calcolato utilizzando un pedigree con informazioni incomplete, è possibile che nel calcolo della depressione da consanguineità si verifichi una sottostima. Per ovviare a questo, alcuni autori hanno proposto di usare il grado di consanguineità individuale (ΔF) invece di F, cosa che sembrerebbe ovviare alle sottostime di cui sopra.
L’effetto della depressione da consanguineità è stato recentemente (Leroy et al, 2014) analizzato tramite una meta-analisi dove sono stati raccolti i risultati della depressione da consanguineità di 57 studi effettuati su 7 diverse specie e riguardanti un totale di 37 caratteri e 5 categorie di caratteri, che andavano dai dati di sopravvivenza ai dati di crescita, a quelli morfologici e a quelli di produzione (latte, lana, carne ecc). Attraverso un modello lineare, sono stati quindi ottenuti i valori attesi dei valori di depressione da consanguineità per ogni aumento unitario percentuale di F dei soggetti. In generale, si è visto che, considerando tutti i caratteri e tutte le specie, l’effetto medio della depressione da consanguineità è pari a 0,137% del valore della media fenotipica del carattere per ogni aumento di 1% F individuale. Quando poi si è calcolata la media per diversi gruppi di caratteri, si è notato che, in generale, i caratteri produttivi sembrano influenzati maggiormente dalla depressione da consanguineità rispetto agli altri gruppi di caratteri.
Per le produzioni di latte, grasso e proteina, le stime espresse in funzione della media sono dell’ordine di -0.367%, -0.249% e -0.225% del valore della media fenotipica dei caratteri, mentre per le cellule somatiche si è arrivati ad una depressione da consanguineità pari allo 0,414% della media. Citando alcuni caratteri di comune interesse zootecnico, si hanno valori pari a -0.191% per la fertilità, +0,322% per la facilità di parto, -0.021% per la durata della gestazione e -0.489% per la sopravvivenza all’età adulta. Per quanto riguarda i caratteri legati al peso, si hanno valori di -0.195% per il peso alla nascita e -0.299% per l’accrescimento.
Impatto Specifico della Consanguineità in Zootecnia
Produzione di latte, grasso e proteina: Diversi studi americani ed europei hanno portato a quantificare, sulla razza Holstein, una diminuzione variabile da 9 a 29 kg per lattazione per un aumento di un punto percentuale di consanguineità del soggetto. Allo stesso modo, sono state quantificate diminuzioni variabili da 0,55 a 1,09 kg di grasso e da 0,67 a 0,96 kg di proteina. Per i caratteri di produzione del latte in genere, le stime di perdita di produzione sono espresse linearmente, cioè hanno lo stesso effetto a qualsiasi livello di consanguineità si trovi l’animale. Una serie di studi ha comunque rilevato che in realtà l’effetto depressivo sulla produzione di latte non è lineare, ma varia come intensità in funzione della classe di consanguineità dell’animale. In effetti, Miglior et al (1992), utilizzando il coefficiente individuale di consanguineità di una popolazione di Jersey canadesi raggruppato in classi di consanguineità e non come effetto continuo, hanno riscontrato che l’effetto depressivo della consanguineità sulla produzione di latte aumentava nelle vacche con consanguineità sopra il 12,5% rispetto alle vacche sotto questa soglia.
Caratteri riproduttivi: Smith et al (1998) hanno riscontrato, per ogni aumento unitario percentuale di consanguineità, una diminuzione della vita produttiva di 5,96 giorni ed un aumento sia della durata dell’interparto (0,31 giorni) che dell’età al primo parto (0,55 giorni). Biffani et al (2002) nella Frisona Italiana hanno riscontrato un aumento dell’intervallo parto-concepimento di 0,31 giorni per ogni 1% di aumento di F. Per quanto riguarda la facilità di parto, McParland et al (2007) hanno riscontrato un aumento di parti distocici fino al 25% di F della vacca, per poi diminuire subito dopo questa soglia, mentre altri autori hanno riportato scarsi effetti sulla distocia, giustificabili anche per il fatto che la facilità di parto viene influenzata dal peso alla nascita del vitello che è minore in vacche consanguinee. Molto interessante è l’effetto della consanguineità che McParland et al (2007) hanno riscontrato sulla sex ratio di vacche consanguinee. È stato notato come vacche con un F fra 5 e 10% avevano, in media, un 1% in più di maschi rispetto alle femmine, mentre sopra il 16% la tendenza si invertiva fino ad arrivare ad un valore atteso del 3% in più di femmine nate per vacche al 25% di consanguineità rispetto a vacche non consanguinee. D’altro canto, si è visto che esiste un aumento di nascite maschili dello 0,1% per ogni punto percentuale di F del vitello: gli autori hanno ipotizzato che quando due animali imparentati si accoppiano, possano aumentare le probabilità che la progenie sia maschile.
Caratteri morfologici: Per quanto riguarda i caratteri morfologici, sia Smith et al (1998) che Biffani et al (2002) che McParland et al (2007) riportano un effetto di aumento dei punteggi morfologici per la maggior parte dei caratteri morfologici legati alla mammella. Come detto, la depressione da inbreeding, impattando negativamente sulle produzioni, ha ovviamente un impatto economico. Appare chiaro, in una situazione zootecnica di avanguardia, che la consanguineità è un aspetto di cui tenere conto. Le Associazioni Nazionali Allevatori e l’Associazione Italiana Allevatori, detentori della tenuta di Libri Genealogici e Registri Anagrafici, hanno gli strumenti, ovvero la tenuta precisa ed aggiornata dei pedigree da cui calcolare il coefficiente di consanguineità dei soggetti e il know-how tecnico ed informatico, e li usano per fornire agli allevatori tutta l’assistenza possibile per quanto riguarda il monitoraggio della consanguineità e le informazioni per gestire la consanguineità in azienda. Nel caso di AIA, essa fornisce questo servizio tramite il suo Ufficio Studi sia per alcune razze in selezione con basso numero di soggetti sotto controllo, sia per le razze del Registro Anagrafico Bovini ed Equidi.
Avanguardia nella Riproduzione: Le Tecniche Assistite
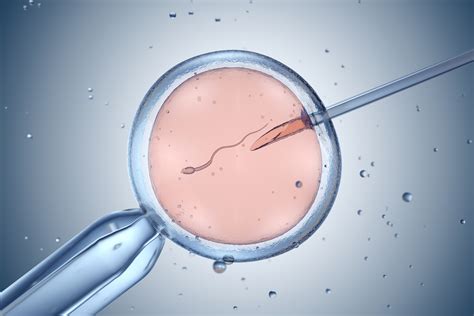
Le tecniche di riproduzione assistita rappresentano un'ulteriore evoluzione nelle strategie riproduttive, spesso con implicazioni dirette sull'allevamento e sulla conservazione delle specie. La fecondazione in vitro (IVF) è un esempio preminente. Nel 1985, Gabriele Schino ha partecipato alla nascita del primo vitello da tecniche IVF e ha lavorato tutta la sua vita alla selezione di ovociti di qualità nelle mucche. La mucca è un buon modello per l'uomo perché ovula una volta al mese e, come la donna, ha un follicolo dominante che crea un uovo.
“Noi abbiamo imparato molto da questo animale”, prosegue Schino, “e possiamo modificare i cicli e avere un accesso maggiore alle ovaie perché non abbiamo i limiti etici che esistono per gli esseri umani, anche se la nostra etica è quella di non fare male agli animali”. La qualità delle uova, infatti, aumenta negli ultimi giorni prima dell'ovulazione ed è un fattore molto sorprendente, che è diverso nel topo, mentre nei bovini, negli umani e nei cavalli è così. Gli ovuli ricevono l’informazione dal follicolo che questo follicolo ovulerà. “Se non c’è questa informazione”, precisa Sirard, “la qualità dell'ovocita non è buona”. Questa è la spiegazione principale per cui il tasso di successo dell’IVF va dal 30 al 40%. La maggior parte delle pazienti non riesce ad ottenere ovociti di qualità a causa dei cambiamenti nei giorni prima dell'ovulazione, che dipendono dal protocollo di stimolazione e dalle condizioni genetiche e ambientali. Quindi, più conosciamo questo modello animale, più possiamo migliorare i trattamenti ormonali nella donna per aumentare la qualità degli ovociti.
tags: #animali #fecondazione #di #gruppo